Le monde de l'horticulture est constamment en quête de nouvelles variétés végétales, alliant esthétisme, résilience et adaptation aux environnements spécifiques. C'est dans cette dynamique qu'un événement marquant a eu lieu à Grand-Champ, en Bretagne : le baptême d'un spécimen de rhododendron inédit, nommé "Grégam", en hommage au nom breton de la commune. Cette création exceptionnelle est le fruit d'un travail méticuleieux d'hybridation, mené par Marc Colombel, expert mondial en rhododendrons, en étroite collaboration avec l'association locale "Balades et Jardins". Ce rhododendron Grégam n'est pas seulement une nouvelle variété ; il représente une avancée significative, étant le premier rhododendron tétraploïde de France, une caractéristique qui ouvre des perspectives fascinantes pour l'avenir de l'espèce.

La Naissance d'une Étoile Horticole : Le Rhododendron Grégam
Le rhododendron Grégam est le résultat de six années d'efforts et de nombreux essais d'hybridation. Selon André Rosnarho, adjoint délégué à l’animation, l'année précédente avait déjà approché la perfection, à l'exception de la couleur. Marc Colombel n'était pas entièrement satisfait, et les conditions météorologiques, notamment le gel, avaient repoussé l'avènement de cette variété d'un an. Cette persévérance a finalement porté ses fruits. La spécificité la plus remarquable du rhododendron Grégam réside dans sa nature tétraploïde, ce qui signifie qu'il possède quatre stocks de chromosomes, contrairement aux deux stocks habituels. Ses deux "parents" sont originaires des États-Unis, conférant au Grégam un patrimoine génétique particulier.
Brighid Goulet-Diskin, surnommée "Bidou" et présidente de l'association Balades et Jardins, désignée marraine du rhododendron Grégam, ne tarit pas d'éloges sur ce spécimen. Elle décrit des fleurs superbes, qui semblent délicatement posées sur les feuilles, avec une épaisseur de pétales bluffante. Le port de l'arbuste est remarquable, lui conférant une allure très élégante. Cette "petite perle" est choyée comme jamais par Pierrick Le Meudec, le responsable des espaces verts de la commune, et son équipe. Le temps clément du week-end de sa présentation a d'ailleurs permis de le découvrir sous son plus bel aspect.
Le rhododendron Grégam a été officiellement baptisé le samedi 11 mai, un événement qui marque une reconnaissance pour Grand-Champ et pour le travail de Marc Colombel. Christian Hays, parrain du rhododendron et historien local, ainsi que Brighid Goulet-Diskin, se chargeront d'inscrire ce spécimen unique à la Royal Horticultural Society, près de Londres. Cet organisme, qui consigne les noms de toutes les nouvelles variétés de plantes au niveau mondial, assurera au Grégam une "superbe reconnaissance internationale".
Les rhododendrons : les points clés pour bien cultiver le rhododendron-arbre à roses - Truffaut
Au-delà de l'Unique : Vers une Troisième Fleur pour Grand-Champ
L'ambition de Grand-Champ ne se limite pas à ce coup médiatique. Comme l'indique le maire, Yves Bleunven, l'objectif est d'améliorer le cadre de vie et le fleurissement de la commune, dans l'espoir d'obtenir une "3e Fleur". Le rhododendron Grégam, symbole de cette démarche d'excellence, restera dans un premier temps dans son bac en plastique, pour être présenté aux habitants de Grand-Champ en le faisant circuler dans toute la commune. Il sera ensuite mis en terre, vraisemblablement dans le parc du Ty Kreiz Ker, où il symbolisera la beauté d'un fleurissement exceptionnel.
Il est important de noter que ce rhododendron unique ne sera pas utilisé à des fins commerciales. Bien qu'il soit possible de tenter de faire des semis ou des greffes, il ne sera jamais reproduit à l'identique. Le rhododendron Grégam est destiné à demeurer un spécimen unique, incarnant l'innovation et l'engagement de Grand-Champ pour la qualité de son environnement.
La Polyploïdisation : Un Processus Clé dans l'Évolution Végétale
La mention de la tétraploïdie du rhododendron Grégam souligne un phénomène biologique fondamental : la polyploïdisation. Ce processus, qui implique la possession de plus de deux jeux de chromosomes dans les cellules d'un organisme, fait partie des phénomènes naturels et spontanés qui contribuent à l'évolution des plantes. La polyploïdie est un moteur important de la spéciation et de l'adaptation chez les végétaux, leur conférant souvent des avantages en termes de taille, de vigueur ou de résistance à certains stress.
Des articles récents rappellent des applications in vitro de la polyploïdisation sur des espèces fruitières et ornementales, en lien avec une meilleure tolérance au stress hydrique. Chez le pommier, par exemple, si la majorité des cultivars sont diploïdes (deux jeux de chromosomes), certains sont triploïdes ou tétraploïdes. La polyploïdisation peut être réalisée in vivo (dans des conditions naturelles) ou in vitro (en laboratoire). Après l'induction de la polyploïdie, l'acclimatation (sortie ex-vitro) et la caractérisation du niveau de ploïdie sont des étapes cruciales pour confirmer la réussite du processus et évaluer les nouvelles propriétés de la plante.
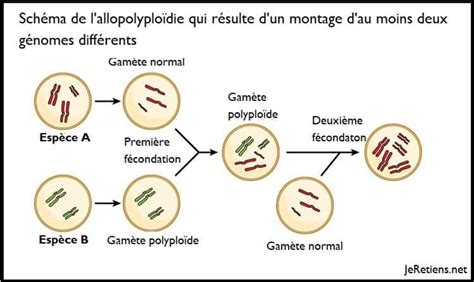
Le Centre R&D de VEGEPOLYS VALLEY propose d'ailleurs d'accompagner les intéressés dans la mise en place de ce type de démarche sur leur propre matériel végétal. La polyploïdisation est une voie prometteuse pour créer de la diversité et améliorer les caractéristiques des plantes cultivées.
L'ADN au Cœur de la Diversité : Comprendre et Utiliser le Patrimoine Génétique des Rhododendrons
L'étude des rhododendrons, qu'il s'agisse de la création de nouvelles variétés comme le Grégam ou de l'analyse de populations sauvages, repose de plus en plus sur l'analyse de leur ADN. Le matériel génétique, en particulier les séquences d'ADN, fournit des informations précieuses sur l'identité, les relations évolutives et les caractéristiques uniques de chaque plante.
Les recherches sur le Rhododendron macrophyllum, le rhododendron de la côte Pacifique, en sont un excellent exemple. Cette espèce, considérée comme unique, s'étend de San Francisco à Victoria et de l'océan Pacifique jusqu'à la crête des Cascades. Sa distribution est cependant très fragmentée, avec de grandes populations continues uniquement dans l'ombre pluviométrique de la chaîne des montagnes Olympiques et dans plusieurs régions de l'Oregon. Les recherches sur cette espèce ont traditionnellement porté sur la couleur des fleurs, qui varie du rose foncé au blanc. Cependant, une étude approfondie a été menée au cours des dix dernières années sur l'ADN des feuilles et des bourgeons floraux du R. macrophyllum.
Les chercheurs impliqués dans ce projet, allant de lycéens à des étudiants diplômés, ont conclu que les populations de rhododendrons de la côte Pacifique contiennent plusieurs types génétiquement distincts, appelés clades. Un clade est un groupe d'organismes censés partager un ancêtre commun non partagé avec d'autres clades. L'analyse du gène de l'intron 4, à partir d'échantillons représentant vingt plantes prélevées à divers endroits, a permis de séparer les gènes en quatre catégories distinctes.
Les données initiales, issues de la thèse d'Amy Denton en 1997, suggéraient que certains clades étaient géographiquement localisés. Par exemple, le clade 1, trouvé en Californie, en Oregon et à Washington, était généralement situé près du niveau de la mer, tandis que les clades 3 et 4 avaient tendance à être présents dans des sites d'altitude plus élevée à l'intérieur des terres. Bien que cette corrélation ne fût pas parfaite et que le nombre total d'échantillons n'était que de 30, une tendance nette a été observée, invitant à des études supplémentaires.
L'Éducation Scientifique et la Recherche Participative
En 2003, une collaboration fructueuse s'est établie entre Laura Sugden, professeure de sciences à la West Seattle High School (WSHS), et Ben Hall, professeur à l'Université de Washington. Des expériences sur l'ADN des rhododendrons ont été réalisées par les étudiants dans leur salle de classe, avec du matériel et des équipements fournis par le Science Education Partnership du Fred Hutchinson Cancer Center de Seattle. L'objectif était d'échantillonner des populations de R. macrophyllum qui n'avaient pas été étudiées dans la recherche d'Amy Denton.
En 2003-2004, la classe a collecté des spécimens à trois endroits : le Mountaineers Theater près de Bremerton, WA, le mont Mitchell (au sud du mont St. Helens) et Cypress Island, près d'Anacortes, WA. Laura Sugden a souligné l'impact positif de ces expériences sur ses élèves : "West Seattle High School est une grande école urbaine où un grand nombre d'étudiants n'ont pas eu beaucoup d'expériences en plein air. En fait, pour certains d'entre eux, ces voyages de collecte ont été parmi leurs premières expériences avec la nature. Tout projet scolaire avec une expérience pratique augmente le niveau d'excitation et d'enthousiasme."
Hien Lu, une étudiante ayant participé à toutes les sorties, a témoigné de l'enrichissement de ces expériences, malgré les défis rencontrés, comme une randonnée pluvieuse et difficile sur Cypress Island. Les séquences d'ADN des échantillons collectés par la WSHS au mont Mitchell et Cypress Island, ainsi que des échantillons de Shawnigan Lake, près de Victoria, C. B., recueillis par Don Whittle, ont parfaitement complété les données d'Amy Denton. Les échantillons de Cypress Island et Shawnigan Lake appartenaient au clade 1 (maritime), tandis que l'échantillon du mont Mitchell était du clade 2, qui semble présent dans toute la région. Ces résultats ont confirmé l'existence de différences génétiques régionales, soulignant la nécessité d'une étude approfondie.
Les rhododendrons : les points clés pour bien cultiver le rhododendron-arbre à roses - Truffaut
Les voyages de collecte et les analyses d'ADN par la classe de Laura Sugden se sont poursuivis l'année suivante. Un programme d'échange étudiant entre l'Université de Washington et l'Université du Sichuan à Chengdu, en Chine, a également intégré cette recherche. Les étudiants chinois, intéressés par l'utilisation de la génétique et de l'analyse d'ADN pour mesurer la diversité végétale, ont trouvé surprenant d'apprendre que le R. macrophyllum était considéré comme une seule espèce, étant donné les différences qu'ils observaient.
En 2004/2005, une collecte à grande échelle a été réalisée le long de la branche canadienne de la rivière Skagit, entre le lac Ross et Rhododendron Flat, sous la direction du professeur Gerry Allen de l'Université de Victoria. Les rhododendrons de Skagit étaient des plantes spectaculairement saines, pleines de bourgeons floraux qui ont été récoltés et conservés dans du gel de silice pour l'extraction d'ADN. Zhenxiang, une étudiante en échange de Sichuan, a ensuite travaillé sur le projet pendant neuf mois. Les résultats ont confirmé la structure de la population de R. macrophyllum. Toutes les copies du gène séquencé (appelé intron 4) appartenaient à l'un des quatre clades. Tous les sites des basses terres du Puget contenaient principalement le clade 1, avec quelques traces du clade 2, et les rhododendrons du haut Skagit appartenaient principalement aux clades 2 et 4, avec un peu de clade 1.
Ces travaux ont été présentés en février 2005 par Xi, Liang, Hall, Sugden, et les lycéens Hien Lu et Olivia Sengsi à un groupe d'experts, membres du Western North America Rhododendron Species Project (WNARSP), ainsi qu'à d'éminents scientifiques.
Explorations Géographiques et Vérification d'Hypothèses
Pour tester l'hypothèse d'une distribution ancestrale maritime, de nouvelles collectes ont été effectuées le long de la côte centrale de l'Oregon. On s'attendait à ce que ces populations aient une forte teneur en gènes du clade 1, si l'hypothèse était correcte. Des expéditions ont été organisées au lac Rhododendron sur l'île de Vancouver, à Wind River et à Florence, en Oregon. Les échantillons obtenus ont renforcé la base de données, et ceux de Florence ont comblé une lacune majeure dans la couverture géographique. Des échantillons supplémentaires ont été envoyés de l'Oregon et de la Californie. L'analyse d'ADN exceptionnellement productive de Zhenxiang en juin et début juillet a permis de déterminer le clade d'ADN pour chacun de ces nouveaux échantillons.

Les données cartographiées (Figure 2) révèlent que la répartition des clades du R. macrophyllum est nettement non uniforme sur son aire de répartition naturelle. La concentration de rouge sur les sites côtiers indique que le clade 1 est généralement trouvé près de l'eau salée, des endroits qui tendent à avoir les températures estivales et hivernales les moins extrêmes. Beaucoup de ces sites se trouvent dans des zones à faible précipitation (ombre pluviométrique). Le clade 3, indiqué en violet, est le type principal trouvé dans les Cascades centrales de l'Oregon. Copper Creek à Washington est le seul autre site où il a été trouvé en abondance.
L'Histoire Écologique et Climatique du Nord-Ouest Américain
L'élaboration d'une hypothèse pour expliquer ces données ADN a nécessité l'entrelacement de découvertes issues de la paléobotanique, de la géologie, de la science atmosphérique, de l'évolution moléculaire et de la systématique végétale. Les différences dans la composition de l'ADN à différents sites suggèrent que les ancêtres de ces populations se sont séparés il y a longtemps et ont suivi des chemins de migration différents. Après leur séparation reproductive, leurs ADN ont divergé.
Des impressions fossilisées de feuilles de rhododendron datant du milieu de l'Éocène (il y a 50 millions d'années [mya]) ont été découvertes à Republic, Washington. Le mélange général de la flore fossile à cet endroit, à Princeton, C.-B., et sur un site de l'Idaho, indique clairement la présence d'une forêt de haute terre, avec des niveaux d'humidité modérés dans l'intérieur du Nord-Ouest pendant et après l'Éocène. Des spécimens fossiles de feuilles et de pollen suggèrent que, tout au long du Miocène (24 à 5 mya), les rhododendrons étaient présents dans l'est de Washington et d'autres parties du Nord-Ouest.
Cependant, on ne trouve aucune trace de R. macrophyllum sur les pentes occidentales des montagnes Olympiques ou des Cascades de Washington au nord du mont Rainier. De même, on ne trouve pas de R. macrophyllum à l'est des Cascades ou dans l'Okanagan canadien. Ces observations indiquent des changements dramatiques dans l'aire de répartition du R. macrophyllum entre le Miocène et le présent. Les données paléobotaniques suggèrent que le retrait du R. macrophyllum de ces régions était dû à la nécessité d'une survie en milieu plus sec, car les pentes occidentales sont devenues trop humides et l'est de Washington a ensuite connu des étés très secs.
Les rhododendrons ont pu survivre sur les crêtes de certaines montagnes des Cascades qui étaient relativement sèches et, à partir de là, ils ont pu migrer vers l'ouest. Cela pourrait expliquer la prévalence des gènes du clade 4 sur la rivière Skagit supérieure en Colombie-Britannique, dans les montagnes du sud de l'Oregon et dans la gorge du Columbia. Pendant le Pléistocène, les glaciers ont dû soumettre les rhododendrons survivants du Nord-Ouest Pacifique à des épreuves encore plus grandes. La majeure partie des zones d'ombre pluviométrique de la moitié ouest du bassin du Puget a été recouverte par une calotte glaciaire de centaines à des milliers de pieds d'épaisseur il y a seulement 16 000 ans, lors de la glaciation de Fraser.
Pourtant, le R. macrophyllum prospère maintenant dans ces zones. Les indications sont que les ancêtres de ces rhododendrons ont persisté pendant le maximum glaciaire dans un petit refuge sur la péninsule Olympique, puis ont repeuplé les péninsules Olympique et Kitsap. En effet, la péninsule Olympique et la péninsule Kitsap, les îles Whidbey et Cypress et le sud de l'île de Vancouver ont des populations de R. macrophyllum très similaires génétiquement. Presque toutes les plantes échantillonnées dans cette vaste zone appartiennent au clade 1.
De même, les grandes inondations du lac Missoula, résultant de la rupture de barrages de glace successifs qui retenaient la rivière Clark Fork, ont été responsables de l'inondation et de l'envasement de la vallée de la Willamette. Le R. macrophyllum est réapparu plus tard dans la vallée de la Willamette et dans certaines parties de l'Oregon. L'histoire écologique et climatique du Nord-Ouest peut en effet fournir une explication à la distribution actuelle des clades 1, 2, 3 et 4 dans toute la région.
Applications Pratiques de la Recherche sur l'ADN des Rhododendrons
La recherche sur l'ADN des rhododendrons, bien que fondamentale, a des conséquences pratiques. Par exemple, les populations de R. macrophyllum autour du canal Hood et des îles voisines, ainsi que dans les Cascades centrales de l'Oregon, sont relativement homogènes génétiquement. En revanche, celles autour de Florence, en Oregon, sont très mélangées génétiquement, incluant divers hybrides entre trois types ancestraux différents (clades 1, 2 et 4).
Parce qu'il existe des différences génétiques entre les populations de différents endroits et parce que de nombreuses populations ont résisté à la sélection dans diverses conditions défavorables, il en découle que des combinaisons de gènes très favorables peuvent être présentes à certains endroits. Ces gènes confèrent une résistance à divers types de stress environnementaux, comme la capacité à résister aux basses températures hivernales (populations du haut Skagit et des Cascades centrales de l'Oregon), à pousser dans des conditions très humides (lac Rhododendron, Wind River), ou à survivre à la sécheresse estivale (péninsule Olympique, canal Hood), pour n'en nommer que quelques-unes.
Ces découvertes ont des implications importantes pour la sélection et la culture des rhododendrons. Les éleveurs pourraient envisager d'établir des jardins d'essai de R. macrophyllum à plusieurs endroits, en choisissant des plantes de divers sites à tester en fonction de leur constitution génétique. Les variétés "Ana", une obtention bretonne, illustre parfaitement cette approche. Bien adaptée aux conditions océaniques, elle forme un arbuste persistant au port arrondi et compact, avec une croissance régulière. Son feuillage vert olive soutenu et sa floraison rose corail lumineux en avril-mai en font un sujet de choix. Sa rusticité correcte (environ -12°C) la rend adaptée à la majorité des jardins de l'ouest, à condition de respecter ses besoins en sol acide et en humidité régulière. Le RHODODENDRON 'Ana' est une preuve de l'importance de sélectionner des variétés adaptées à des conditions spécifiques.

Créer de la Diversité : Les Marqueurs Moléculaires et la Sélection Végétale
La recherche de caractères nouveaux est constante chez les espèces cultivées. Elle concerne les obtenteurs dans le cadre de programmes de création variétale et les producteurs qui recherchent des variations spontanées. Ces variants peuvent provenir de mutations, un mécanisme crucial, naturel et spontané de l'évolution des espèces. Cependant, en raison du caractère ponctuel des mutations, il peut s'avérer difficile d'obtenir un individu uniforme et stable.
C'est là que les marqueurs moléculaires entrent en jeu. Ils permettent une caractérisation rapide, précise, précoce, fiable et économique du matériel végétal. Le Centre R&D de VEGEPOLYS VALLEY, par exemple, réalise le sexage (mâle/femelle) via l'analyse de l'ADN de l'arbre (tests PCR), à partir de très petits échantillons (une feuille par arbre). Cela permet, par exemple, de garantir un Ginkgo mâle, évitant les nuisances liées aux fruits des femelles en zone urbaine.
Les marqueurs moléculaires sont devenus des outils indispensables pour la sélection assistée par marqueurs. Lorsqu'un marqueur moléculaire lié à un caractère d'intérêt est mis en évidence, il peut être utilisé pour la sélection. Cela permet aux hybrideurs d'optimiser leurs programmes de création variétale.
Dans le cas du pommier, la pollinisation en vergers est un enjeu majeur. Le pommier est allogame, présentant une auto-incompatibilité pollinique qui empêche l'auto-fertilisation via un mécanisme de reconnaissance pollen-pistil. Ce système est contrôlé par le locus "S", avec une série d'allèles d'auto-incompatibilité (S1, S2, S3… S47 par exemple). La connaissance de ces allèles S est cruciale pour l'hybrideur. Si les géniteurs sont complètement incompatibles, aucun hybride ne sera obtenu. Avec des parents semi-compatibles, seulement 50 % des croisements attendus seront réussis.
Le gène MdPG1 (Polygalacturonase 1, chromosome 10) joue un rôle essentiel dans les processus de désassemblage des parois cellulaires lors de la maturation et dans la texture du fruit du pommier. Il est déjà utilisé par les programmes de création variétale pour la qualité de la pomme. De même, les Malus floribunda sont des pollinisateurs bien connus, mais cette espèce est aussi la source de caractères d'intérêt comme la résistance à la tavelure (locus Rvi6 ou Vf) et au puceron cendré.
Les analyses d'ADN ne se limitent pas à l'identification variétale ou au sexage. Elles sont également utilisées pour le phytodiagnostic, c'est-à-dire l'identification des agents pathogènes présents sur les cultures. Le service phytodiagnostic du CENTRE R&D de VEGEPOLYS VALLEY offre une identification ciblée ou globale des pathogènes (bactéries, champignons, virus, nématodes) par séquençage, et même par microscopie électronique ou RT-PCR pour les virus. Ces analyses sont possibles à partir de feuilles, de bourgeons, de bois, et même de fruits, permettant des diagnostics en toute saison.
Les Stimulateurs de Défense des Plantes : Une Nouvelle Approche pour la Protection
En parallèle de la sélection variétale, la recherche explore des moyens d'améliorer la résistance des plantes aux maladies. Les stimulateurs de défense des plantes (SDP) représentent une approche prometteuse. Chez la vigne, une dizaine de produits de protection à effet stimulateur de défense sont recommandés, à base de chito-oligosaccharides avec oligogalacturonides (COS-OGA), d'extraits de Cerevisane, de Phosphonates de potassium ou de Bacillus amyloliquefaciens. D'autres produits SDP, comme le chitosan, l'acibenzolar-S-méthyl, la laminarine ou le fenugrec, sont commercialisés sur toutes cultures.
Ces produits agissent en déclenchant des mécanismes de défense complexes dans la plante. En quelques heures, les flux ioniques sont modifiés, avec une importance particulière du calcium pour l'activation des protéines kinases. Des molécules hormonales clés, telles que l'acide salicylique (SA), l'acide jasmonique (JA) et l'éthylène (ET), sont ensuite impliquées dans la régulation des gènes de défense. Par exemple, l'acibenzolar-S-méthyl induit la résistance systémique acquise (SAR) de la plante via la voie du SA. En quelques jours, les protéines PR s'accumulent. Les phytoalexines, comme les stilbènes chez la vigne, sont également produites en réponse tardive. Ces polyphénols, dérivés du trans-resvératrol, ont une activité antifongique directe et un rôle antioxydant.
Le Centre R&D de VEGEPOLYS VALLEY étudie l'expression des gènes et les profils de production des stilbènes dans le cadre du projet collaboratif MISTIC, afin de mieux comprendre et optimiser ces mécanismes de défense. Pour perfectionner leurs analyses, ils ont couplé la technologie de l'expression quantitative des gènes par qPCR avec les cartes FTA®, qui permettent de récolter des extraits foliaires au champ. Ces cartes, utilisées pour collecter des échantillons sanguins, sont un moyen facile de collecte et de transport des acides nucléiques. Après broyage des feuilles et dépôt des extraits sur la carte, les acides nucléiques sont capturés et stabilisés sur la membrane. Cette méthode innovante permet de mesurer les niveaux d'induction des gènes de défense ciblés et de visualiser les modalités présentant une induction.
Le Monde Fascinant des Rhododendrons
Les rhododendrons forment une grande famille d'arbustes, la plupart du temps persistants, adaptés à de nombreuses régions de France grâce à leur bonne résistance au froid. Ce sont des plantes acidophiles, ne supportant pas le calcaire, qui peut entraîner des chloroses et la mort de la plante à court terme. L'origine botanique des rhododendrons commercialisés est essentiellement l'Asie du Sud-Est, même si certaines espèces poussent naturellement dans d'autres régions. Cette diversité, de l'Himalaya au Japon, apporte une grande disparité dans les formes et les couleurs.
Dans une approche horticole, on sépare souvent les rhododendrons des azalées, bien que botaniquement, toutes ces plantes appartiennent au genre Rhododendron. Parmi les azalées, on distingue les persistantes (Azalea japonica) et les caduques (Azalea mollis). Les Azalea japonica, par leur développement modéré et leurs formes, sont parfaites pour les petits massifs variés. Elles s'adaptent à toutes les expositions, du plein soleil aux espaces plus ombragés, et supportent des pH relativement élevés (jusqu'à 6.7/6.8). Elles présentent une grande diversité de formes (prostrées ou érigées), de tailles de feuilles et de couleurs de fleurs en trompette, regroupées en corymbe.

La plantation du rhododendron 'Ana', par exemple, se fait de préférence à l'automne ou au printemps, en évitant les périodes de gel ou de forte chaleur. Il nécessite un sol acide, humifère et bien drainé, de type terre de bruyère. Il est conseillé de creuser un trou de 4 à 5 fois la taille de la motte, d'ajouter de la corne broyée ou du BOCHEVO, puis de tasser la terre ameublie autour de la plante. L'exposition idéale est la mi-ombre, sous une lumière filtrée ou à l'abri d'un arbre léger. Le rhododendron peut tolérer le soleil en Bretagne si le sol reste frais. Un "petit truc" pour la plantation est de passer toute la périphérie de la motte au jet d'eau après l'avoir sortie du pot, afin de dégager les petites radicelles et de les orienter correctement.
L'entretien du rhododendron 'Ana' est limité si les conditions de plantation sont respectées. L'arrosage doit être suivi les premières années et en période sèche, car la plante redoute les sols qui se dessèchent. La taille n'est pas indispensable ; on peut simplement supprimer les fleurs fanées pour favoriser la formation de nouveaux boutons. Un apport annuel de terre de bruyère ou de compost de feuilles au printemps est bénéfique. Le paillage est important pour maintenir un sol frais et acide. Ce rhododendron est globalement facile, à condition d'éviter les sols calcaires et mal drainés.
Le rhododendron 'Ana' s'intègre naturellement dans un massif de terre de bruyère, en association avec d'autres plantes de sol acide, comme des azalées caduques (AZALEA mollis ‘Gibraltar’), des rhododendrons plus compacts (RHODODENDRON ‘Percy Wiseman’), des arbustes persistants (PIERIS japonica ‘Mountain Fire’, Kalmia latifolia ‘Pimwheel’) et des vivaces adaptées aux sols acides (HEUCHERA ‘Dark Secret’, DRYOPTERIS erythrosora ou GAULTHERIA procumbens). Ces associations permettent de créer des massifs équilibrés et évolutifs au fil des saisons, dans une ambiance douce et légèrement ombragée. Le RHODODENDRON 'Ana' supporte le climat maritime et le vent, et est une plante à feuillage persistant, ce qui en fait un atout précieux pour les jardins bretons.
En définitive, du spécimen unique Grégam aux vastes populations sauvages du R. macrophyllum, l'étude des rhododendrons révèle une richesse insoupçonnée, tant sur le plan génétique qu'écologique. Ces recherches, menées par des experts, des étudiants et des passionnés, contribuent non seulement à la création de nouvelles variétés, mais aussi à une meilleure compréhension de l'évolution et de l'adaptation des plantes.
tags: #adn #en #commun #avec #rhododendron