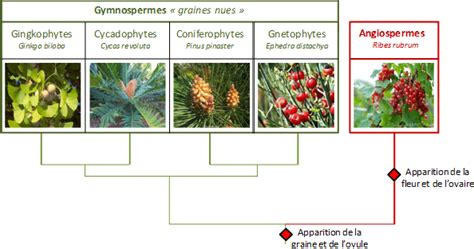
Le règne végétal, d’une diversité extraordinaire, est peuplé de formes de vie adaptées à des environnements variés. Parmi elles, les plantes à graines, ou Spermaphytes, représentent un jalon évolutif majeur, ayant conquis la plupart des écosystèmes terrestres. Ce groupe se divise principalement en deux grandes catégories : les Gymnospermes et les Angiospermes. Si elles partagent la capacité de produire des graines, leur stratégie reproductive et la structure de leurs organes diffèrent considérablement. Les Angiospermes, avec leurs fleurs complexes et leurs fruits protecteurs, dominent aujourd'hui la plupart des écosystèmes terrestres, tandis que les Gymnospermes, un groupe plus ancien et souvent associé aux conifères, présentent des caractéristiques reproductives distinctes. Cet article se propose d'explorer en profondeur les organes reproducteurs et leur fonctionnement chez ces deux groupes de plantes, mettant en lumière les adaptations qui ont forgé leur succès évolutif.
Les Angiospermes : L'Apogée de la Fleur et du Fruit
Les Angiospermes, dont le nom signifie « graine dans un récipient » en grec, représentent la plus grande partie des espèces végétales terrestres, avec une estimation allant de 250 000 à 300 000 espèces. Elles dominent les paysages naturels terrestres tropicaux et tempérés, comme la savane ou la forêt. Leur succès évolutif est attribué à la grande nouveauté de leur système reproducteur : la fleur, qui contient des carpelles se transformant en fruits, constituant la principale synapomorphie du groupe. La reproduction sexuée des Angiospermes permet de générer de nouveaux individus génétiquement différents de leurs parents, grâce aux brassages génétiques réalisés par la méiose et la fécondation.
La Fleur d'Angiosperme : Une Merveille d'Ingénierie Biologique
Une fleur est issue du développement d’un bourgeon floral terminal ou latéral. Elle est portée par un pédoncule inséré à l’aisselle d’une feuille (la bractée), et l’ensemble est relié à un rameau. Les caractères morphologiques, le nombre et les relations entre les pièces florales peuvent varier d’une espèce à l’autre.
Les fleurs de lis (espèces du genre Lilium) sont particulièrement faciles à étudier du fait de leurs grandes dimensions, permettant de réaliser aisément coupes et dissections florales. Elles ont de fait beaucoup été utilisées pour des coupes histologiques qui ont historiquement permis de comprendre la reproduction des Angiospermes. L'observation attentive d’une fleur épanouie de lis permet de décrire assez bien sa structure. Les pièces florales forment quatre verticilles, c’est-à-dire quatre cercles concentriques de pièces similaires. Elles s’attachent au réceptacle floral, un renflement de l’extrémité du pédoncule floral.
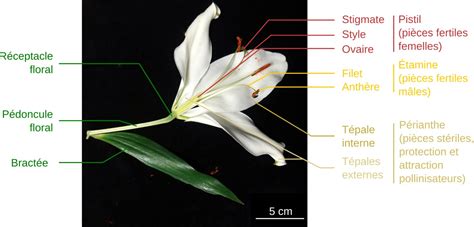
Les Pièces Florales Stériles : Protection et Attraction
Le périanthe est un ensemble de pièces florales stériles protégeant les organes reproducteurs. Il se compose généralement de deux verticilles :
- Le Calice : Il est l'ensemble des sépales.
- La Corolle : Elle est formée par l'ensemble des pétales.
Chez le lis, les pièces florales des deux premiers verticilles ne sont pas distinguables morphologiquement et sont appelées tépales. L'ensemble du calice et de la corolle forme le périanthe. Il arrive que le calice et la corolle soient difficiles à différencier et, dans ce cas, on parle de tépales à la place des pétales et des sépales et le périanthe est nommé périgone. Les tépales sont de grandes structures foliacées, colorées et odorantes chez le lis, attirant les pollinisateurs.
Les Organes Reproducteurs Mâles : L'Androcée
Le troisième verticille correspond à l’androcée, appareil reproducteur mâle de la fleur, formé par les étamines. Chez le lis, il y a un verticille de six étamines. Chaque étamine est constituée d’un filet surmonté d’une anthère.
Formation du pollen
L'Anthère et la Production de Pollen
La partie fertile d’une étamine est l’anthère. Pour en décrire la structure, il faut réaliser des coupes dans le bouton floral avant l’épanouissement de la fleur. L’anthère est formée de deux paires de sacs polliniques (sporanges mâles) à l’intérieur desquels se forment les grains de pollen. À maturité, les sacs polliniques s’ouvrent au niveau de la zone de déhiscence entre chaque paire et libèrent le pollen. En coupe, la symétrie bilatérale de l’anthère évoque la silhouette d’un papillon. Chacune des quatre « ailes » est un sac pollinique qui protège la sporogenèse et la production des grains de pollen avant de les libérer. L’assise mécanique présente une paroi lignifiée et agit comme un ressort au moment de l’épanouissement.
Chez les Angiospermes, deux types de spores sont produites : on parle d’hétérosporie. Les grosses spores sont qualifiées de mégaspores (ou macrospores) et les petites de microspores. Les microspores sont produites par les microsporanges, sur une microsporophylle (l’étamine, une autre feuille modifiée) et donnent par mitoses les microgamétophytes.
La formation des grains de pollen commence par la prolifération de cellules mères de spores, diploïdes, contenues dans les sacs polliniques. Ces cellules mères des spores s’entourent d’une paroi de callose et subissent une méiose qui produit quatre spores mâles haploïdes, processus qui marque le passage dans la génération gamétophytique du cycle de vie. La paroi de callose est ensuite dégradée et chaque spore se divise par mitose, pour donner la cellule végétative et la cellule générative, cette dernière étant incluse dans le cytoplasme de la cellule végétative. En plus de leur rôle nutritif, les cellules du tapis s’immiscent entre les spores et synthétisent la sporopollénine de l’exine.
Le Grain de Pollen : Le Gamétophyte Mâle
Les grains de pollen sont facilement observables en les prélevant sur les anthères des fleurs épanouies et en les déposant entre lame et lamelle. Il s’agit de structures d’environ 100 µm, ovoïdes et présentant une paroi ornementée. Les gouttelettes jaunes à la surface (le pollenkitt), associées aux reliefs en réseau, permettent aux grains de pollen de mieux adhérer aux pollinisateurs.
Le grain de pollen est le gamétophyte mâle. Il est constitué de seulement deux cellules haploïdes contenues dans une paroi complexe et très résistante. Le gamétophyte mâle, tout comme le gamétophyte femelle, est donc très réduit, comme chez toutes les plantes à graines actuelles, mais contrairement à ce que l’on peut observer chez les Embryophytes dites « à spores libres » (mousses et fougères). La cellule végétative occupe la majorité du volume. Après la pollinisation, elle permettra la formation du tube pollinique, qui transporte les gamètes. À l’intérieur de la cellule végétative se trouve la cellule générative. Les cellules sont très déshydratées et contiennent des réserves (lipides, glucides notamment), ce qui est typique de formes de vie ralentie. La partie interne (intine) de la paroi protectrice du grain de pollen est pectocellulosique, et la partie externe (exine) est composée de sporopollénine, le composé biologique le plus résistant connu, assurant une protection mécanique et chimique. Cette cellule générative produit deux gamètes mâles par mitose.
Les Organes Reproducteurs Femelles : Le Gynécée
Le quatrième verticille est le gynécée, appareil reproducteur femelle de la fleur, formé de carpelles renfermant un ou plusieurs ovules. Chez le lis, on observe un verticille de trois carpelles soudés par la longueur, qui forment le pistil. Le nombre de lobes du stigmate, ici trois, permet de supposer que le gynécée est formé de trois carpelles. Les carpelles peuvent être libres ou soudés (partiellement ou entièrement).
Le Pistil et l'Ovaire
La partie fertile du pistil est son ovaire. Sa structure à différentes échelles microscopiques peut être observée en réalisant des coupes transversales. L’ovaire de lis est formé de trois carpelles soudés. Il forme une sorte d’hexagone lobé et plus ou moins régulier. Chaque carpelle contient une loge carpellaire le long de laquelle sont disposés les ovules, par groupes de deux. Durant leur formation, les ovules sont protégés de la dessiccation et des agressions physiques par l’épiderme et l’épaisse paroi des carpelles. Ces mêmes structures protègent, après la fécondation, les graines issues des ovules.
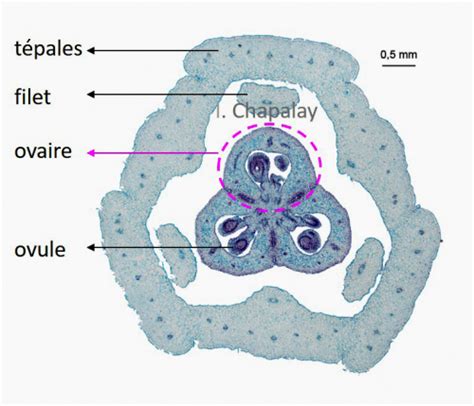
L'Ovule et le Sac Embryonnaire
Les ovules des Angiospermes sont des structures pluricellulaires à l’intérieur desquels est produit le gamète femelle, l’oosphère. Ils sont situés dans les ovaires et sont reliés à la paroi du carpelle par le funicule. L’ovule est constitué, de l’extérieur vers l’intérieur, de deux téguments protecteurs (caractéristique des Angiospermes, les graines des Gymnospermes n’en possédant qu’un seul), du nucelle et du sac embryonnaire qui lui-même contient le gamète femelle.
Le sac embryonnaire correspond au gamétophyte femelle. Il est très réduit, constitué de seulement sept cellules avec huit noyaux chez le lis, comme chez de nombreuses espèces d’Angiospermes. L’oosphère, le gamète femelle (haploïde), se trouve du côté du micropyle avec deux synergides. Une cellule centrale comporte deux noyaux polaires. Trois antipodes se trouvent à l’opposé du micropyle (côté chalaze). Le funicule relie le corps de l’ovule (au niveau de la chalaze) au carpelle (au niveau du placenta).
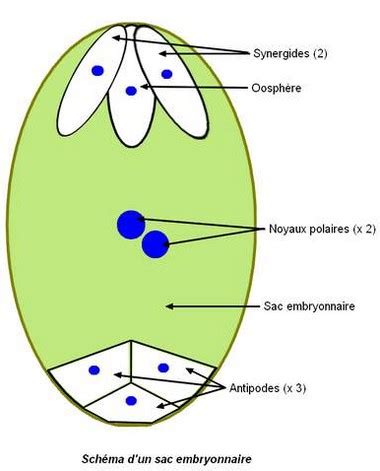
La formation du sac embryonnaire commence par la méiose de la cellule mère des spores, contenue dans le nucelle. Après méiose, quatre noyaux haploïdes sont produits au sein d’une même cellule (il n’y a pas de cytocinèse après les divisions nucléaires). Trois des noyaux haploïdes fusionnent en un noyau triploïde. Après deux mitoses, toujours sans cytocinèse, huit noyaux sont obtenus. Enfin, l’ensemble se cellularise pour former un sac embryonnaire. Le noyau haploïde, côté micropyle, est à l’origine de l’oosphère (gamète femelle), des deux synergides et d’un des noyaux polaires.
La Double Fécondation : Une Spécificité des Angiospermes
Chez les Angiospermes, la fécondation est double. Un des gamètes mâles du grain de pollen fusionne avec l’oosphère pour former le zygote diploïde, qui donnera l’embryon. L'autre gamète mâle fusionne avec les deux noyaux polaires de la cellule centrale pour former le noyau de l'albumen, qui, en se développant, donnera l'endosperme, un tissu nourricier triploïde qui servira de réserve pour l'embryon en développement.
La Graine et le Fruit : Dispersion et Protection
Après la fécondation, les ovules se transforment en graines et les carpelles en fruit.
Formation du pollen
La Graine d'Angiosperme
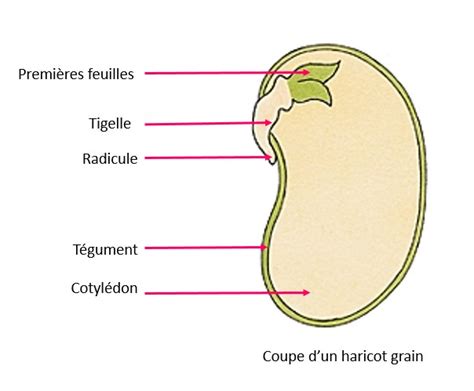
La graine contient l'embryon diploïde, formé à partir de la fusion d'un gamète mâle et de l'oosphère. Le zygote diploïde se développe pour former un embryon multicellulaire. Les réserves nutritives, qui alimenteront l'embryon lorsque la graine sera parvenue à maturité, peuvent être stockées dans l'endosperme ou dans les cotylédons de l'embryon, qui sont souvent très développés.
Les téguments de l'ovule deviennent le tégument de la graine, composé de deux couches : la testa (externe) et le tegmen (interne). Ce tégument protecteur s'épaissit pour devenir une enveloppe rigide et imperméable qui se déchire au moment de la germination. L'embryon occupe la partie centrale de la graine. Sa taille varie considérablement d'une essence à l'autre. Il est composé d'une radicule, d'une tigelle, d'une gemmule et de deux cotylédons chez les dicotylédones, ou un seul chez les monocotylédones. Des cotylédons supplémentaires ou absents peuvent être observés.
La durée de maturation des graines varie considérablement d'une espèce à l'autre. Par exemple, chez Eucalyptus marginata, elle est de 5 à 6 mois, tandis qu'elle est de dix à seize mois chez Eucalyptus camaldulensis. Chez Pinus merkusii, il faut environ 200 jours avant qu'elle soit parfaitement mûre.
Le Fruit d'Angiosperme
Le fruit est une structure qui dérive de l'ovaire de la fleur après fécondation. La paroi de l'ovaire s'épaissit pour former le péricarpe, qui est la partie du fruit qui contient la graine. Le péricarpe peut être sec, semi-charnu ou charnu, et sa structure varie considérablement. On distingue trois couches : l'épicarpe (peau externe), le mésocarpe (chair) et l'endocarpe (noyau ou partie interne).
Les fruits sont des dispositifs de protection et de dissémination des graines. La dissémination des graines peut se faire de diverses manières :
- Anémochorie (par le vent) : Les fruits secs (samares de Fraxinus, Betula) comportent des ailes ou des poils servant à prolonger le vol. Les graines peuvent être transportées sur des centaines de mètres.
- Zoochorie (par les animaux) : Les fruits charnus ou semi-charnus (drupes de Gmelina, Azadirachta, Mangifera) ont un goût particulier qui attire les oiseaux et les animaux frugivores. Les graines sont consommées et déposées dans les fèces à une distance considérable du lieu d'ingestion. La digestion amollit le tégument, facilitant la germination.
- Autochorie (par la plante elle-même) : Certains fruits secs s'ouvrent par déhiscence, libérant les graines à proximité de la plante mère. Par exemple, une capsule fend le long de deux sutures, libérant les graines.
La dormance des graines est un mécanisme adaptatif qui retarde la germination, permettant aux graines de survivre à des conditions défavorables. La levée de dormance peut être provoquée par des facteurs environnementaux comme le froid (stratification) ou le passage dans le tube digestif d'un animal.
Les Gymnospermes : Des Graines Nues et des Cônes Évolués
Les Gymnospermes sont des plantes faisant partie d'un sous-embranchement paraphylétique des Spermaphytes. Le terme gymnosperme signifie « graine nue », ce qui permet de définir cette classe végétale dans un sens large, renfermant un certain nombre de phylums arrivés à un même stade évolutif. Les Gymnospermes s'opposent aux Angiospermes par leur graine non enfermée dans un fruit. Ce sont des intermédiaires entre les cryptogames vasculaires (Ptéridophytes) et les Angiospermes.
Les Gymnospermes sont des arbres ou des arbustes à feuillage persistant, à l'exception de quelques genres à feuillage décidu (mélèze, cyprès chauve de Virginie). Ils sont bien représentés par les Conifères (sapins, épicéas, mélèzes, séquoias, pins, genévriers, cyprès, thuyas), dont on connaît la valeur économique et ornementale.
Les Organes Reproducteurs : Les Cônes
Les organes reproducteurs sont regroupés en fleurs encore peu évoluées chez les gymnospermes. C'est à l'intérieur de certains organes de ces fleurs que les gamétophytes se développeront. Les conifères sont des espèces dioïques ou monoïques, mais leurs fleurs sont toujours unisexuées. L'appareil reproducteur des Gymnospermes est constitué d'inflorescences de type monopodial (grappe ou épi) et unisexuées. L'organisation florale est variée, et on dénombre de très fréquents états préfloraux, mais les fleurs typiques sont rares du fait que le périanthe ne soit présent qu'exceptionnellement.
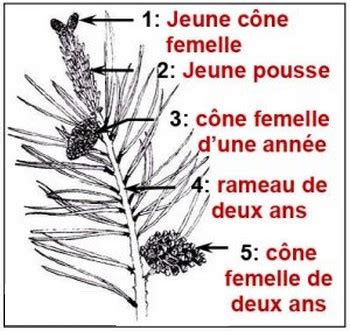
L'Organe Reproducteur Femelle : Le Cône Femelle
Les fleurs femelles de conifère sont regroupées en cônes plus ou moins allongés (ex : la pomme de pin). Le cône est un ensemble de fleurs femelles ou inflorescence femelle. Les fleurs s'insèrent en spirale sur l'axe du cône et chacune est constituée d'une bractée (écaille réduite) à l'aisselle de laquelle se trouve une écaille ovulifère portant le plus souvent deux ovules.
Formation du pollen
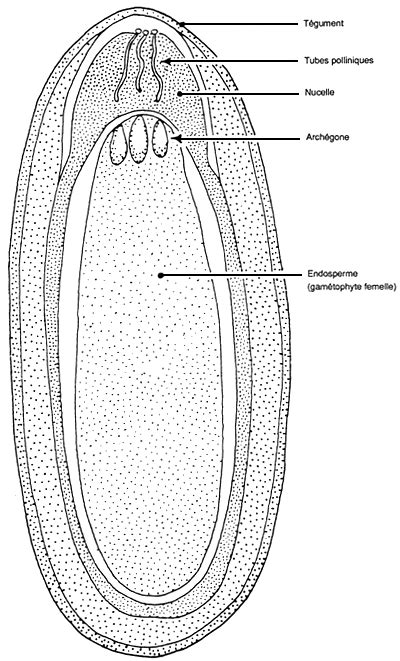
L'Ovule Nu
Les ovules sont nus sur l'écaille ovulifère, ou feuille carpellaire, directement accessibles au pollen. Il n'y a pas de chambre pollinique comme chez l'ovule des préspermaphytes. L'ovule est toujours entouré par un seul tégument protecteur. Ce tégument renferme le nucelle ou macrosporange dans lequel se forme après méiose quatre macrospores. Trois dégénèrent, la macrospore restante se développe au sein du nucelle par mitose pour donner le gamétophyte femelle ou "endosperme". L'endosperme, bien que réduit à quelques centaines de cellules, possède encore deux archégones rudimentaires qui se différencient au pôle apical ou pôle micropylaire. Une seule oosphère par ovule sera fécondée.
L'Organe Reproducteur Mâle : Le Cône Mâle
Les fleurs mâles forment de petits cônes constitués uniquement d'étamines insérées en spirale sur un petit axe court. Ces étamines, aux formes diverses, sont de petites écailles crochues chez le pin. Elles portent sur leur face inférieure deux sacs polliniques.
Les Sacs Polliniques et la Production de Pollen
À l'intérieur de ces sacs polliniques a lieu la méiose pour donner des microspores qui ne sont pas disséminées, mais qui se développent par mitose pour donner les gamétophytes mâles ou grains de pollen, très réduits (quelques cellules) qui eux seront disséminés. Le pollen se forme, en général, par cloisonnements simultanés, généralement pourvu de sacs aérifères, sauf chez certaines familles (Araucariaceae, Cephalotaxaceae, Taxodiaceae et Cupressaceae). Il est presque toujours pluricellulaire, rarement syncytial. La déhiscence des sacs polliniques se fait par un exothécium. Chaque grain de pollen renferme deux gamètes mâles nus (noyaux gamétiques non vermiculaires).
La Fécondation Simple et Différée
Chez les Gymnospermes, la fécondation est simple, et souvent très décalée par rapport à la pollinisation. Par exemple, chez les Pinus, un cône femelle met trois ans à mûrir après la pollinisation. Lorsque le grain de pollen atteint le sac embryonnaire, il libère deux gamètes mâles dont l'un fusionne avec l'oosphère. Le produit de cette union forme, en se développant, le nouvel embryon diploïde. L'autre gamète mâle dégénère.
La Graine de Gymnosperme : Une Graine Nue
Après fécondation, les cellules prothalliennes prolifèrent et se gorgent de réserves. Le gamétophyte femelle massif et cellulaire constitue le tissu nourricier, appelé aussi endosperme primaire, chez les gymnospermes. Ce tissu haploïde fournit des réserves nutritives à l'embryon. Cet usage du terme endosperme primaire a été désapprouvé par certains, préférant le terme de prothalle femelle.
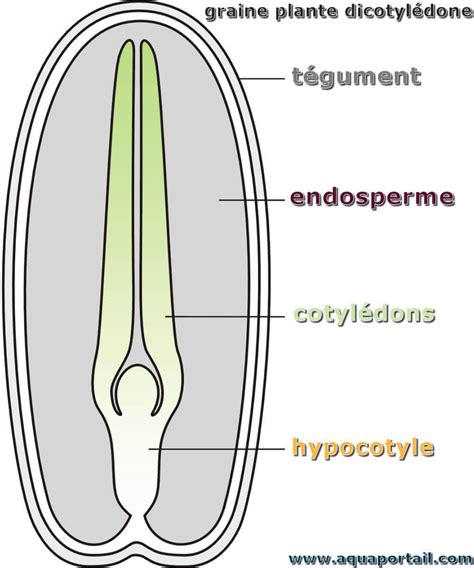
La graine de Gymnosperme est caractérisée par son ovule unitégumenté. Le tégument de l'ovule devient le tégument de la graine. Contrairement aux Angiospermes, la graine n'est pas enfermée dans un fruit.
L'embryon, occupant la partie centrale de la graine, est composé d'une radicule, d'une tigelle et de cotylédons. Le nombre de cotylédons varie considérablement d'une essence à l'autre ; par exemple, le Pinus comporte en général de deux à quinze cotylédons, tandis que la plupart des arbres angiospermes n'en comportent que deux. Un seul embryon par graine parvient à maturité dans la grande majorité des cas.
Les Gymnospermes présentent divers mécanismes de dissémination des graines :
- Anémochorie : Les graines peuvent être ailées, comme chez certains genres de Pinus, facilitant la dispersion par le vent.
- Zoochorie : Chez certains genres, comme Taxus ou Juniperus, le tégument de la graine est entouré d'une excroissance charnue et vive, l'arille, qui attire les oiseaux et les mammifères.
La période entre la floraison et la maturité des graines varie considérablement. Par exemple, elle est de 21 à 24 mois chez Araucaria hunsteinii.
Les cônes des Gymnospermes s'ouvrent généralement sous l'effet de la dessiccation des écailles pour libérer les graines. Cependant, chez certains genres comme Pinus, il existe des pins “à cônes fermés” (sérotonie), comme Pinus contorta, qui ne s'ouvrent et ne libèrent les graines qu'à la suite d'un événement perturbateur, tel que des feux de forêt occasionnels.
Similarités et Divergences Évolutives
Le cycle de vie des lis (espèces du genre Lilium) est un cycle typique des Embryophytes, c'est-à-dire où une génération sporophytique, diploïde et produisant les spores, alterne avec une génération gamétophytique, haploïde et produisant des gamètes. Cette alternance est une caractéristique fondamentale partagée par les Angiospermes et les Gymnospermes. Cependant, les Angiospermes présentent des adaptations plus poussées pour la protection de l'embryon et la dissémination des graines, ce qui a contribué à leur succès évolutif.
Le terme de Gymnosperme couvre une série de formes intermédiaires, pour beaucoup d'organes, entre les Ptéridophytes et les Angiospermes. Si, par exemple, on étudie l'appareil femelle, on voit l'archégone complexe des Pteridophytes se simplifier chez les Gymnospermes et encore plus chez les Angiospermes. Ce caractère intermédiaire apparaît quand on étudie les divers aspects de ces plantes, mais il faut toujours songer qu'il s'agit d'évolutions parallèles.
En résumé, la distinction fondamentale entre Angiospermes et Gymnospermes réside dans la présence du fruit pour les premières, qui protège la graine, et l'absence de cette protection chez les secondes. Cette différence a des implications majeures sur leur stratégie reproductive, leur interaction avec l'environnement et leur répartition géographique actuelle.
tags: #organe #reproducteur #de #certaines #plantes