La reproduction sexuée des plantes à fleurs, ou angiospermes, est un processus complexe et fascinant, au cœur duquel se trouve le stigmate. Cet organe, partie intégrante du pistil, joue un rôle déterminant dans la réception et la reconnaissance des grains de pollen, essentiels à la perpétuation de l'espèce. Pour le rhododendron, qu'il s'agisse de l'espèce ornementale ou de l'envahisseur comme le Rhododendron ponticum, la compréhension du rôle du stigmate et du transport du pollen est primordiale, tant pour l'horticulture que pour la gestion écologique.
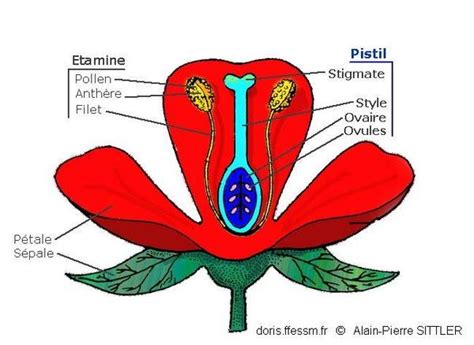
L'Architecture du Pistil et la Fonction du Stigmate
En botanique, le stigmate est défini comme l'extrémité d'un carpelle, ou de plusieurs carpelles soudés, formant le pistil d'une fleur. Il constitue, avec le style et l'ovaire, le pistil, lui-même composant une partie du gynécée, c'est-à-dire l'ensemble des organes reproducteurs femelles d'une plante à fleurs. Le stigmate est la partie distale du style ou stylodia. Sa structure est spécialisée pour la capture du pollen. Il est composé de papilles stigmatiques, des cellules dont la fonction première est de capturer les grains de pollen. Ces cellules peuvent être localisées uniquement à l'extrémité du style ou, chez certaines espèces, notamment celles pollinisées par le vent (anémophilie), elles peuvent couvrir une surface beaucoup plus large, augmentant ainsi l'efficacité de la capture.
Le rôle fondamental du stigmate est de recevoir le pollen. C'est sur cette surface que se produit la germination du grain de pollen, étape indispensable à la fécondation. Pour optimiser la capture, le stigmate est souvent collant en surface et adapté par diverses caractéristiques morphologiques telles que des poils, des languettes ou des reliefs.
La Réception et la Reconnaissance du Pollen
Le pollen peut parvenir au stigmate de diverses manières : transporté par l'air (anémophilie), par des insectes ou d'autres animaux visiteurs (pollinisation biotique), ou plus rarement, par l'eau environnante (hydrophilie). Une fois déposé, le stigmate ne se contente pas d'accueillir le pollen ; il joue un rôle actif dans sa reconnaissance et sa viabilité.
Il a été démontré que le stigmate facilite la réhydratation du pollen, un processus crucial car le pollen est généralement très déshydraté lorsqu'il quitte l'anthère. Cette réhydratation est essentielle pour favoriser la germination du tube pollinique. De plus, le stigmate assure une bonne adhérence des grains de pollen d'espèces adaptées, permettant ainsi au processus de se poursuivre.
Un aspect fascinant de la fonction stigmatique est sa capacité à discriminer le pollen. Le stigmate peut jouer un rôle actif dans la reconnaissance du pollen et dans certaines réactions d'auto-incompatibilité. Ces mécanismes permettent à la plante de rejeter le pollen de plantes identiques ou génétiquement proches, favorisant ainsi le brassage génétique et l'augmentation de la diversité de l'espèce. Un stigmate, en effet, ne réagira pas au pollen d'une autre espèce. Bien que la forme physique des grains de pollen soit très particulière, suggérant une reconnaissance géométrique, d'autres stimulations et réponses biochimiques jouent également un rôle crucial.
L'Évolution de la Pollinisation : Des Spores aux Grains de Pollen Spécialisés
L'histoire évolutive des plantes terrestres, qui sont apparues il y a 410 millions d'années, témoigne d'une libération progressive de la contrainte de l'eau pour la reproduction, d'une facilitation du brassage génétique, d'une sécurisation de la fécondation et du développement de l'embryon, ainsi que de la mise en place de mécanismes assurant la survie et la dispersion de la descendance sexuée.
Chez les fougères, par exemple, ce sont des spores et non des grains de pollen qui sont dispersées. Ces spores, plus fragiles, donnent naissance, dans des conditions favorables, à un prothalle qui produit des gamètes. La fécondation, où les gamètes mâles nagent, ne peut se dérouler sans eau.
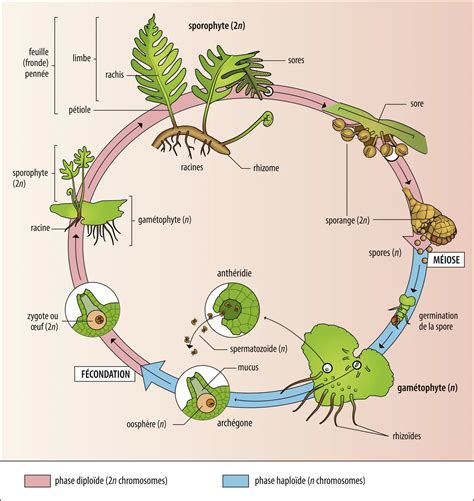
Les Gymnospermes, apparues il y a 260 millions d'années, marquent une étape cruciale avec l'apparition du pollen et de la graine, qui sécurisent la fécondation et facilitent le brassage génétique. Chez les Gymnospermes, la pollinisation est généralement assurée par le vent, comme en témoignent les nuages de grains de pollen chez les pins.
Enfin, les Angiospermes, apparues il y a environ 130 millions d'années, représentent le groupe de plantes le plus diversifié. Caractérisées par des graines enfermées dans des ovaires et une double fécondation, elles ont développé des relations riches et variées avec les insectes. Le pollen chez les Angiospermes, surtout lorsqu'il est transporté par les insectes, est de grande taille et possède une exine (paroi externe) ornementée pour faciliter sa fixation sur le corps des insectes. Il est produit par des étamines souvent placées à l'intérieur de fleurs colorées et bien visibles, produisant du nectar.
Le Rhododendron : Un Cas d'Étude de la Pollinisation par les Insectes
Le Rhododendron, en particulier le Rhododendron ponticum, originaire de Turquie et de la péninsule ibérique, est un excellent exemple de plante dont la reproduction est fortement dépendante des insectes pollinisateurs. Le Rhododendron des parcs se multiplie par reproduction sexuée, et ses belles inflorescences roses ou violettes sont un attrait majeur pour les pollinisateurs.
Insectes Pollinisateurs et Efficacité
La pollinisation du Rhododendron ponticum peut être assurée par plusieurs espèces d'insectes, selon le pays colonisé. En Irlande, il a été observé que les bourdons (Bombus spp.) participent activement à la pollinisation des fleurs de Rhododendron ponticum. Des études ont même révélé que ces bourdons transportaient plus de pollen en Irlande qu'en Espagne, suggérant des adaptations ou des interactions différentes dans les zones d'introduction par rapport aux habitats natifs.
Pour d'autres espèces de rhododendrons, comme le Rhododendron ferrugineum, diverses espèces d'insectes visiteurs ont été enregistrées, notamment les Hyménoptères (abeilles, bourdons), les Diptères (mouches), les Coléoptères et les Lépidoptères (papillons). Les abeilles domestiques et les bourdons sont souvent les insectes les plus fréquents et efficaces. Les mouches muscides sont également considérées comme d'importants pollinisateurs en raison de leur taux de visite relativement élevé. En revanche, les syrphes, les fourmis (Formicidae) et les coléoptères se sont avérés inefficaces pour le transport du pollen, tandis que l'efficacité des lépidoptères et des empididés était négligeable.
Dispersion du Pollen et Implications Génétiques
Une expérience de marquage par fluorescence a révélé que la dispersion du pollen chez le Rhododendron ferrugineum est restreinte (généralement entre 0 et 2 mètres) dans un peuplement dense, et qu'elle diminue de manière leptocurtique. Cette restriction peut favoriser l'auto-pollinisation géitonogame (transfert de pollen entre différentes fleurs de la même plante), ce qui pourrait expliquer la forte relation génétique entre les individus observée dans les études génétiques du Rhododendron ferrugineum. Pour de nombreuses plantes alpines, qui sont majoritairement allogames (pollinisation croisée), le succès reproductif dépend fortement de l'efficacité des pollinisateurs. La compréhension de la manière dont les pollinisateurs transfèrent le pollen d'une fleur à l'autre est d'un grand intérêt pour comprendre la structure génétique des populations végétales.
Quel est rôle de la pollinisation ?
Les Interactions Complexes : Pollinisateurs, Maladies et Espèces Invasives
Les relations entre les plantes à fleurs et les insectes sont anciennes, remontant aux Angiospermes ancestrales où la pollinisation était déjà assurée par des coléoptères se nourrissant de pollen. Ces relations sont devenues complexes et diversifiées, offrant gîte et couvert aux insectes tout en assurant la pollinisation de 80% des plantes à fleurs. Les adaptations morphologiques réciproques entre l'animal et la plante conduisent parfois à des relations spécifiques, souvent qualifiées de co-évolution.
Cependant, ces interactions peuvent aussi avoir des répercussions écologiques importantes, notamment avec l'introduction d'espèces invasives comme le Rhododendron ponticum.
Le Rhododendron ponticum : Une Espèce Invasive
Le Rhododendron ponticum est une espèce invasive qui, bien qu'encore discrète dans les milieux naturels de France et de Bretagne, est dotée de capacités de colonisation rapide. En Grande-Bretagne, l'invasion est particulièrement importante. Les nombreux jardins publics et privés constituent des sources extrêmement importantes de propagules de cette espèce. Le principal motif de ces introductions multiples est l'horticulture, la plante étant très appréciée pour ses belles inflorescences.
L'impact écologique du Rhododendron ponticum est significatif. Il crée un ombrage important qui limite drastiquement le développement des autres végétaux. De plus, il est soupçonné d'être un réservoir important pour le développement de champignons du genre Phytophthora, comme le fatal Phytophthora cinnamomi. En Angleterre, des bilans économiques de l'invasion par le Rhododendron pontique ont été réalisés pour estimer le coût des mesures de gestion, révélant des dépenses considérables (670 924 livres sterling pour gérer cette espèce).
Malgré ses nuisances potentielles, le Rhododendron ponticum ne fait pas encore l'objet de mesures de contrôle de ses populations dans toutes les régions, comme en Bretagne où il semble bien s'adapter. La gestion de cette plante est complexe : la coupe simple des troncs ne suffit pas en raison des capacités de rejet à partir des souches. L'application d'un herbicide sur les souches, directement après la coupe ou quelques années après, est nécessaire. Cependant, le traitement aux herbicides ne doit pas être utilisé à la légère.
La Cicadelle du Rhododendron : Un Vecteur de Maladie
Outre les interactions avec les pollinisateurs, le rhododendron est également la cible d'autres insectes, certains étant vecteurs de maladies. La cicadelle du rhododendron, Graphocephala fennahi, originaire d'Amérique du Nord, est un exemple. Cet insecte, aux teintes vives d'orange et de vert, mesure moins d'un centimètre et est apparu en Grande-Bretagne dans les années 1930, puis en Europe vers les années 1970, se répandant lentement mais sûrement. Il est considéré comme un ravageur et une espèce invasive, introduit via le commerce des rhododendrons.
Les cicadelles adultes sont visibles entre juillet et novembre, période durant laquelle la ponte a lieu directement dans les écailles des bourgeons floraux en formation. Cette cicadelle ne donne naissance qu'à une seule génération par an. Les larves, blanchâtres à vert jaune, restent sur la face inférieure des feuilles d'avril à juillet-août, se nourrissant de la sève des arbustes. Ce ponctionnement affaiblit la plante, et la salive des larves contient une toxine qui bloque la circulation de la sève, entraînant dessèchement et nécrose des feuilles. Le miellat excrété par les larves favorise également la formation de fumagine, qui ralentit la photosynthèse.
Le danger principal de la cicadelle du rhododendron réside dans sa capacité à propager les spores d'un champignon, le Pycnostysanus azaleae, en même temps qu'elle pond. Ce champignon provoque la maladie du "Bud blast" ou "bouton noir", caractérisée par le noircissement des boutons floraux. Ces derniers brunissent d'abord, puis se couvrent de minuscules "clous" noirs, qui sont les fructifications du champignon. Les rameaux peuvent prendre une teinte grisâtre et mourir si plusieurs boutons sont touchés.
La lutte contre la cicadelle est essentielle pour prévenir la maladie. Elle implique des mesures préventives et des méthodes de lutte biologique ou écologique, comme l'utilisation d'huile de colza (attention aux butineurs), de pyrèthre naturel, de kaolinite, de savon noir dilué, ou encore de purins de rhubarbe ou d'ortie, et d'huile de neem. Des conditions de culture adaptées, un sol bien drainé, et une limitation des engrais azotés qui attirent les ravageurs sont également importantes. La suppression des boutons noircis dès que possible et leur destruction sont cruciales pour éviter l'expansion de la maladie.
Diversité des Mécanismes de Pollinisation
La nature a développé une incroyable diversité de mécanismes de pollinisation, chacun adapté aux spécificités de la plante et de ses pollinisateurs.
Les Iris : Une Stratégie d'Allopollinisation
Les Iris, plantes herbacées géophytes à rhizome ou bulbes, sont des exemples intéressants de pollinisation entomophile favorisant l'allopollinisation. Leurs fleurs hermaphrodites, régulières et trimères, possèdent des sépales réfléchis et des pétales dressés. Les trois étamines sont libres et situées à la base des pétales. L'ovaire est infère, et le style se prolonge en trois lobes stigmatiques, souvent développés en forme de limbe et constitués de deux lèvres superposées cachant entre elles des papilles stigmatiques.
Lors de la pollinisation, les abeilles sont attirées par le nectar sécrété par des glandes à la base des sépales. En passant entre les sépales et les lobes stigmatiques, elles frottent leur dos, portant du pollen d'une autre fleur, sur les stigmates. Après avoir prélevé le nectar et en ressortant de la fleur, elles prélèvent du pollen de la fleur qu'elles viennent de butiner. Le pollen déposé sur les stigmates développe alors un long tube pollinique qui transporte les gamètes mâles vers les ovules. Cette méthode favorise la fécondation entre deux individus différents et limite l'auto-fécondation, même si les fleurs sont hermaphrodites, augmentant ainsi le brassage génétique et la diversité de l'espèce.
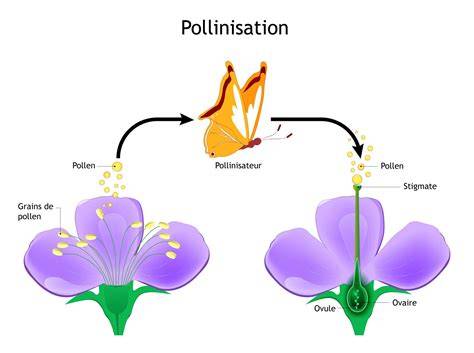
Les Salvia : Un Mécanisme de Levier
Chez les sauges (Salvia), les fleurs colorées et de grande taille attirent les insectes pollinisateurs. L'insecte, en entrant dans la corolle, fait basculer les étamines qui déposent le pollen sur son dos. Ce mécanisme de levier assure un dépôt précis du pollen sur le pollinisateur.
Les Orchidées : Mimétisme et Leurre Sexuel
Les orchidées sont maîtresses dans l'art d'attirer les pollinisateurs grâce à divers facteurs : production de nectar (genres Spiranthes et Orchis), rôle important de l'éperon, mimétisme avec les plantes à nectar (genre Orchis), et même leurre sexuel (genre Ophrys). Chez Ophrys scolopax, par exemple, le labelle imite la forme de l'insecte pollinisateur. Le mâle, adulte avant la femelle, est attiré par un bouquet d'odeurs similaire à la phéromone sexuelle de la femelle, participant ainsi à la pollinisation. La morphologie de la fleur permet ensuite la fixation du pollen sur le corps de l'animal, son transport, puis son dépôt sur le stigmate du pistil. L'hybridation interspécifique est même possible chez certaines orchidées, comme entre l'orchis singe et l'orchis homme pendu, en raison d'un pollinisateur commun.
Les Aracées : Piège et Protogynie
Les Aracées, telles qu'Arum italicum, attirent les insectes pollinisateurs par une odeur nauséabonde émise par l'inflorescence (spadice enveloppé d'une spathe). Elles présentent une protogynie, où les fleurs femelles situées vers le bas de l'inflorescence sont fertiles avant les fleurs mâles. Des fleurs stériles régulent le passage des insectes. Chez Arum italicum, les insectes chargés de pollen sont bloqués par des fleurs stériles dans l'ampoule de la spathe pour permettre la pollinisation et la fécondation des fleurs femelles fertiles. Une fois cela fait, les insectes peuvent remonter vers les fleurs mâles devenues fertiles, se charger à nouveau de pollen, et quand les appendices des fleurs mâles stériles se flétrissent, ils peuvent quitter la spathe pour polliniser une autre plante (allopollinisation).
Les Moracées : Un Mutualisme Complexe chez les Figuiers
La pollinisation des figuiers (Moracées) est un mutualisme particulier et complexe. Leurs inflorescences sont protogynes. Les larves de pollinisateurs se développent au sein de fleurs femelles stériles qui se transforment en galle. À maturité, les insectes s'envolent et se chargent de pollen au contact des fleurs mâles fertiles avant de quitter l'inflorescence. Quand les insectes femelles vont pondre dans de jeunes inflorescences de figuier, seules les fleurs femelles sont fertiles et reçoivent le pollen que les insectes avaient prélevé dans l'inflorescence précédente. Ce mutualisme implique aussi des insectes parasites des pollinisateurs et des fourmis prédatrices, attirées par ces derniers ainsi que par des messages chimiques olfactifs.
Pollinisation Anémophile : Le Cas des Graminées
À l'inverse des plantes pollinisées par les insectes, chez les plantes à fleurs pollinisées par le vent (anémophiles), le pollen est de petite taille, à exine lisse, et produit dans des anthères pendantes avec de longs filets, au sein de fleurs réduites et discrètes. Chez les graminées, par exemple, les fleurs sont discrètes (sépales et pétales absents) et les anthères sont fixées au filet par leur milieu, facilitant leur balancement par le vent et la libération d'un pollen de petite taille et à paroi lisse.
Le Safran : Un Stigmate Précieux

L'espèce Crocus sativus (Iridaceae) est cultivée pour ses stigmates, qui sont récoltés et séchés pour produire le safran. Ce dernier est une denrée d'un prix exorbitant, entre 3000 et 25000 Euros le kilo, ce qui s'explique par la nécessité de près de 140 000 fleurs pour obtenir 5 kilos de stigmates, qui se réduisent à un kilo une fois séchés. Chaque fleur de Crocus sativus, qui s'épanouit d'octobre à novembre, porte un style grêle divisé en trois branches épaisses et rouges : les stigmates. La cueillette est journalière et très exigeante en main-d'œuvre. Le safran est utilisé comme épice, colorant alimentaire, et en médecine pour ses propriétés stimulantes et emménagogues.
D'autres stigmates, comme ceux du maïs (Zea mays), appelés "soies de maïs" ou "cheveux de maïs", sont également utilisés dans le domaine médicinal. Ces exemples illustrent la diversité des fonctions et de la valeur des stigmates dans le règne végétal.
Conclusion Partielle
Le stigmate, bien plus qu'une simple extrémité, est un acteur central de la reproduction des plantes à fleurs. Sa morphologie et ses fonctions biochimiques sont finement adaptées pour la capture, la réhydratation et la reconnaissance du pollen, assurant ainsi la fécondation et le maintien de la diversité génétique. Des relations complexes avec les insectes pollinisateurs aux défis posés par les espèces invasives et les maladies, l'étude du stigmate et du grain de pollen offre un aperçu profond des mécanismes qui façonnent la vie végétale sur Terre.