Le système cardiovasculaire des poissons, bien que partageant des principes fondamentaux avec celui des autres vertébrés, présente des adaptations uniques, notamment la présence du bulbe artériel. Cette structure, essentielle à la régulation du flux sanguin, se distingue par ses caractéristiques spécifiques et son rôle crucial dans la circulation des poissons. Cette étude détaillée se propose d'explorer le bulbe artériel chez les poissons, en mettant en lumière sa définition, son fonctionnement, ses distinctions par rapport à d'autres structures cardiaques et son importance dans le contexte plus large de la physiologie des poissons.
Le Bulbe Artériel : Une Chambre Cardiaque Distincte
Le bulbe artériel, également connu sous le nom de bulbus arteriosus, représente une particularité anatomique remarquable dans le cœur de certains poissons. Il s'agit d'une locution masculine qui désigne la chambre la plus antérieure du cœur chez les Téléostéens et les Cyclostomes. Contrairement à son équivalent chez d'autres poissons, le bulbe cardiaque, le bulbe artériel se caractérise par l'absence de valvules et une nature non contractile. Il constitue une partie dilatée de l'aorte située juste en avant du cœur, d'où proviennent les arcs aortiques chez les embryons de vertébrés et chez l'adulte de nombreux vertébrés inférieurs.
Dans le système circulatoire des poissons, le bulbe artériel est une chambre en forme de poire. Cette configuration lui permet de fonctionner comme un condensateur, une fonction cruciale pour maintenir un flux sanguin continu vers les arcs branchiaux. C'est la quatrième des chambres connectées en série au sein du cœur des poissons, succédant au sinus veineux, à l'oreillette et au ventricule. Le sang, après avoir traversé ces cavités, est pompé dans l'aorte abdominale via le bulbe artériel. Sa paroi est dotée d'une structure à trois couches, ce qui lui confère la capacité d'agir comme une installation de stockage intermédiaire et de soutenir ainsi un flux sanguin régulier vers les branchies.
Il est intéressant de noter qu'une structure comparable au bulbe artériel est traversée pendant les phases de développement du cœur humain et est également appelée bulbus cordis. Celui-ci se trouve ventralement au ventricule primitif après que le cœur en développement a pris sa forme en S. Ensemble, le bulbus cordis et le ventricule primitif forment les ventricules du cœur ainsi structuré. Les parois adjacentes du bulbus cordis et du ventricule se rapprochent, fusionnent et finissent par disparaître, de sorte que le bulbus cordis communique désormais librement avec le ventricule droit, tandis que la jonction du bulbe avec le tronc artériel est amenée ventralement dans le canal auriculaire.
Distinction entre Bulbe Artériel et Bulbe Cardiaque
La terminologie peut parfois prêter à confusion, il est donc essentiel de bien distinguer le bulbe artériel du bulbe cardiaque. Le bulbe cardiaque, ou bulbus cordis en anatomie des vertébrés, identifie la chambre la plus antérieure du cœur des poissons autres que les Téléostéens et des embryons de vertébrés. À la différence du bulbe artériel, les parois du bulbe cardiaque sont épaisses, musculeuses et comportent de nombreuses valvules. Il est en liaison avec l'aorte ventrale par l'intermédiaire du tronc artériel. Le système cardiaque d'un poisson est généralement divisé en quatre parties, dont le bulbe cardiaque peut être une composante animée.
Chez l'homme et les mammifères, le bulbe cardiaque est un ventricule primitif qui apparaît après que le cœur en développement a pris sa forme en "S". Le bulbus cordis et le ventricule primitif donnent naissance aux ventricules du cœur formé. L'extrémité supérieure du bulbus cordis est également appelée conotroncus. Par l'excroissance de la cloison ventriculaire, le bulbe cardiaque est en grande partie séparé du ventricule gauche, mais reste une partie intégrante du ventricule droit, dont il forme l'infundibulum. Autrement dit, le bulbus cordis est une région du cœur fœtal primitif qui apparaît à la fin de la troisième semaine du développement du cœur humain et animal.
Les synonymes du bulbe artériel incluent bulbe aortique, bulbus arteriosus et bulbus cordis (dans un contexte embryonnaire). Son antonyme est le bulbe cardiaque, soulignant la différence fonctionnelle et structurelle entre ces deux entités.
Le Cœur des Poissons et le Circuit Sanguin
Respirer avec des branchies cycle 4
Le cœur est l'organe principal du système circulatoire sanguin chez les poissons. Il est l'organe qui envoie le sang aux artères qui irriguent les différentes branchies, ce sont les artères branchiales afférentes. Le sang repart des branchies par des artères branchiales efférentes qui vont distribuer le sang oxygéné aux organes. Il est important de noter que le cœur des poissons n'est pas double comme chez les mammifères (pas de cœur droit veineux, pas de cœur gauche artériel) ; il est uniquement veineux. Le sang arrive par le sinus veineux, passe par une cavité appelée atrium (équivalent de l'oreillette), puis pénètre dans le ventricule. Ce dernier possède une paroi musculaire importante. Lorsque le ventricule se contracte, il envoie le sang dans le bulbe artériel, puis dans l'aorte ventrale.
Le système circulatoire des poissons est clos et comporte une pompe cardiaque. Le circuit sanguin forme une boucle unique : le sang va du cœur aux branchies, puis part irriguer le reste du corps avant de revenir au cœur, à l'intérieur duquel ne circule que du sang veineux. Chez la plupart des poissons, l'oxygénation du cœur par les coronaires est limitée à l'épicarde, la couche externe du cœur, et l'irrigation des tissus cardiaques se fait principalement par le sang contenu au sein de la cavité cardiaque. Le cœur des actinoptérygiens se compose de quatre cavités : le sinus veineux qui collecte le sang veineux et le dirige vers l'atrium ; l'atrium, poche musculaire à sens unique qui dirige le sang vers le ventricule ; le ventricule cardiaque, poche musculaire qui effectue le travail de pompage cardiaque ; le bulbe cardiaque (ou bulbe artériel chez les Téléostéens) qui, relié à l'artère aorte, permet l'éjection du sang vers les branchies. Chez les poissons pulmonés, le cloisonnement du cœur est analogue à celui des tétrapodes pulmonés.
L'étude de l'évolution des espèces soulève la question de la complexité. Les poissons à nageoires rayonnées, ou actinoptérygiens, constituent la classe de poissons la plus diversifiée de tous les vertébrés, incluant les téléostéens qui représentent 99,8 % des espèces de poissons actuelles. Les biologistes considèrent que le cœur d’un actinoptérygien se compose de quatre cavités : un collecteur - le sinus veineux - dirige le sang veineux vers une poche musculaire à sens unique - l’atrium - qui l'envoie dans une autre poche musculaire effectuant le travail de pompage - le ventricule - et finalement dans un organe qui régule la pression du sang avant éjection vers les branchies, qui peut prendre la forme d'un « bulbe artériel », ou d'un « cône artériel ». Des paléontologues ont remarqué depuis le XIXe siècle cette apparente simplification de la physiologie du cœur des poissons actinoptérygiens, mais ignoraient quand et comment elle avait pu se produire.
Pour répondre à cette question, des chercheurs ont étudié des fossiles de Rhacolepis buccalis, un poisson ayant vécu pendant le Crétacé. Leurs analyses ont révélé un cône artériel de forme cylindrique constitué d'une épaisse paroi musculeuse, dont l’intérieur comportait une série de cinq anneaux hélicoïdaux, interprétés comme les restes fossilisés de composantes fibreuses des valves. Cette observation est interprétée comme une simplification évolutive, Polypterus, un actinoptérygien basal africain, possédant pas moins de neuf rangées de valves dans son cœur, tandis que Rhacolepis buccalis n’en possède que cinq. Certains téléostéens primitifs tels que les tarpons en ont deux, et les téléostéens n’en possèdent qu’une, leur cône artériel ayant pratiquement disparu pour être réduit à la forme d’un bulbe. Les paléontologues avancent que trois événements de simplification se seraient produits, de manière convergente, dans les lignées actinoptérygiennes.
Anatomie Cardiaque et Contraction chez les Poissons
Le cœur des poissons présente des caractéristiques anatomiques distinctes de celles des mammifères. En général, le cœur des poissons est composé de quatre compartiments principaux disposés en série : le sinus veineux, l'oreillette, le ventricule et le cône artériel (ou bulbe artériel chez certaines espèces).
Le sinus veineux est une chambre à paroi mince qui reçoit le sang veineux du corps. L'oreillette est une chambre à paroi mince qui reçoit le sang du sinus veineux et le pompe dans le ventricule. Le ventricule est une chambre à paroi épaisse qui pompe le sang vers les branchies. Enfin, le cône artériel (ou bulbe artériel) est une structure élastique qui amortit les variations de pression et assure un flux sanguin continu vers les branchies.
La contraction cardiaque chez les poissons est un processus complexe qui implique une coordination précise des événements électriques et mécaniques. Les cardiomyocytes, cellules musculaires cardiaques, sont responsables de la contraction du cœur. Ces cellules présentent des jonctions particulières, des jonctions pour la résistance et des desmosomes. Les striations transversales montrent la présence de sarcomères qui sont des agencements particuliers d’actine et de myosine, proche de l’agencement cristallin. On trouvera dans ces cardiomyocytes de grandes quantités de mitochondries, des capillaires sanguins entre deux fibres, des disques intercalaires ou stries scalariformes (ancien terme) qui sont des jonctions entre deux cardiomyocytes. Les parois des cardiomyocytes sont proches mais ne se touchent pas.
Le potentiel d'action cardiaque des poissons est similaire à celui des autres vertébrés, avec une phase de dépolarisation rapide, une phase de plateau et une phase de repolarisation. La phase de plateau est principalement due au calcium. La différence de potentiel transmembranaire se passe de part et d’autre de la membrane. Les premiers ions calcium entrant (extracellulaires) vont aller stimuler la sortie de Ca2+ du réticulum sarcoplasmique.
Le couplage excitation-contraction est le processus par lequel le potentiel d'action cardiaque déclenche la contraction musculaire. Chez les poissons, ce processus implique l'entrée de calcium extracellulaire à travers les canaux calciques voltage-dépendants, ce qui induit la libération de calcium du réticulum sarcoplasmique. L'augmentation de la concentration de calcium intracellulaire active les protéines contractiles, actine et myosine, entraînant la contraction musculaire.
Régulation de la Contraction Cardiaque
La contraction cardiaque chez les poissons est régulée par divers facteurs, notamment le système nerveux autonome, les hormones et les facteurs locaux.
Le système nerveux autonome, composé des branches sympathique et parasympathique, joue un rôle important dans la régulation de la fréquence cardiaque et de la force de contraction. L’action de la noradrénaline se fait sur la contraction du muscle cardiaque. Elle va augmenter cette force de contraction. La noradrénaline se lie sur un récepteur B2 adrénergique membranaire et entraîne la formation d’AMPc. Le calcium participe à l’augmentation de calcium dans le réticulum endoplasmique et permettant une plus forte vague calcique. Il agit aussi en stimulant la pompe, ce qui fait rentrer le calcium dans le réticulum endoplasmique et ce qui raccourcit ainsi la durée du flash (et un relâchement plus rapide). Le cœur présente également des récepteurs muscariniques pour l’acétylcholine. L’acétylcholine va diminuer la force de contraction. Le système orthosympathique agit par un accroissement de la vitesse de contraction et de la force maximale.
Diverses hormones, telles que l'adrénaline et le cortisol, peuvent influencer la contraction cardiaque chez les poissons. Des facteurs locaux, tels que la température et le pH, peuvent également affecter la contraction cardiaque.
Circulation Sanguine et Oxygénation Spécifique
La circulation permet, entre autres, le transport de l’oxygène qui est devenu nécessaire suite à l’accroissement de taille des organismes. Le liquide interstitiel, qui baigne les cellules, est composé de 10,5 litres. La circulation sanguine ne représente qu’une faible proportion des liquides corporels. Pour être efficace, elle a une vitesse importante.
Pour effectuer le calcul d’un volume, on dilue une substance dans le nouveau volume. Pour l’eau totale, on utilise de l’eau marquée au tritium (3H2O). Pour le plasma, on utilise l’albumine. Cette dernière ne quitte pas la circulation sanguine (en réalité, elle ne quitte que très peu ce compartiment). L’albumine utilisée sera marquée radioactivement par de l’iode (radioactif). La circulation est en relation avec l’eau extracellulaire. Chez les mammifères, l’appareil circulatoire est clos mais pas étanche.
Le sang artériel est diffusé dans les capillaires où se situe le transfert de l'oxygène et des aliments aux tissus environnants. Les éléments nutritifs sont absorbés par les intestins, transportés vers le foie et plus tard diffusés dans le sang. Dans les reins, le sang est « purifié » et les déchets rejetés par l'urine. Les veines se rassemblent dans un seul vaisseau avant d'arriver au cœur. La quantité totale du sang est de 1,5 à 3 % du poids du corps, et les tissus musculaires, qui constituent les deux tiers du poids du corps, ne contiennent que 20 % du volume sanguin : les muscles blancs ne sont pas très vascularisés.
L'Échocardiographie : Un Outil d'Exploration Cardiaque
L'échocardiographie, une technique d'imagerie non invasive basée sur les ultrasons, est un outil essentiel pour l'étude du cœur des poissons, bien que son application chez l'homme soit plus répandue. Elle permet de visualiser dynamiquement le cœur, ses cavités et ses valves, ainsi que les principaux vaisseaux. Elle permet également de caractériser les flux sanguins et le déplacement tissulaire grâce au Doppler.
L'échocardiographie utilise des ultrasons pour créer des images du cœur. Une sonde est placée sur le thorax du patient (ou de l'animal), après avoir été enduite d'un gel conducteur pour faciliter le passage des ultrasons à travers la peau. Un tracé ECG est enregistré simultanément pour analyser les structures anatomiques en fonction de la phase du cycle cardiaque.
L'examen échocardiographique est d'abord un examen morphologique dynamique, permettant d'obtenir des coupes des structures cardiaques en 2 dimensions au cours du cycle cardiaque. La technique TM (temps - mouvement) représente le déplacement des structures cardiaques sur une ligne au cours du temps.
L'étude morphologique est couplée à une analyse Doppler, qui permet de mesurer la vitesse de déplacement du sang ou des structures cardiaques au cours du temps. Le Doppler Pulsé (DP) permet de mesurer les vitesses de flux dans une zone spécifique, mais est limité par le phénomène d'aliasing. Le Doppler Continu (DC) permet d'enregistrer des vitesses de flux très élevées, mais ne renseigne pas sur le point d'origine des vitesses. Le Doppler Couleur utilise la technologie du DP avec un affichage des vitesses encodé en couleur, permettant d'identifier l'origine, la direction et le type de flux. Enfin, le Doppler Tissulaire (DTI) permet d'étudier le mouvement des parois myocardiques en se basant sur le décalage de fréquence de l'onde réfléchie sur le tissu myocardique.
Différentes modalités techniques sont utilisées selon les informations qui peuvent être obtenues sur chaque fenêtre d'exploration. La Fenêtre Parasternale Grand Axe (PSGA) permet de mesurer le diamètre du ventricule gauche (VG) en télédiastole et en télésystole, les épaisseurs des parois, et le diamètre de l'aorte ascendante. La Fenêtre Para Sternale Petit Axe (PSPA) permet de visualiser le ventricule droit (VD), le VG, les piliers de la valve mitrale, la valve aortique, l'artère pulmonaire, la valve tricuspide et les oreillettes. La Fenêtre Apicale : Les coupes 4, 2, 3 cavités permettent d'analyser la cinétique segmentaire du VG, la fonction systolique ventriculaire du VG, et de mesurer le volume de l'OG. La Fenêtre Sous-Costale permet de visualiser les 4 cavités cardiaques et un épanchement péricardique.
Respiration et Digestion chez les Poissons
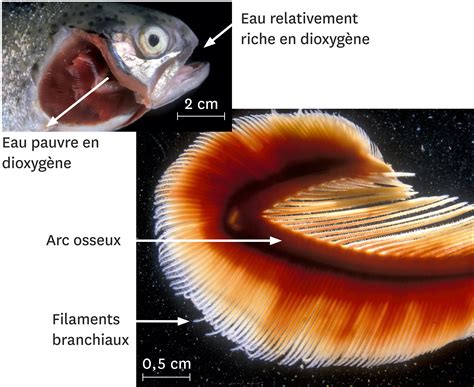
La plupart des poissons pratiquent usuellement la respiration aquatique à travers leurs branchies. Les poissons pompent l'eau (contenant du dioxygène) par la bouche, puis la font circuler sur les branchies. Chez la plupart des poissons cartilagineux (hormis les chimères), les fentes branchiales sont visibles (latéralement chez les requins et les lamproies, ventralement chez les raies). Le sang est aéré dans les branchies : le poisson avale l'eau, les branchies captent l'oxygène de l'eau et l'envoient vers le sang. L'eau traverse les branchies et ressort. Pour respirer, le poisson va fermer alternativement sa bouche et ses opercules. Le courant d'eau produit va irriguer ses branchies. Le sang fixe l'oxygène à travers la fine paroi des branchies et libère le gaz carbonique.
Il est important de noter que l'air contient 200 cm3 d'oxygène par litre, tandis que l'eau est saturée pour 9 cm3 à 5 °C et pour 5 cm3 à 30 °C. Le système respiratoire du poisson doit donc prendre en compte cette énorme différence.
Certains poissons ont développé des adaptations respiratoires remarquables. La peau des anguilles peut être utilisée pour absorber l'oxygène. Les périophthalmes respirent en absorbant l'oxygène à travers la peau, de façon similaire aux grenouilles. Un « organe de respiration annexe » peut être utilisé pour extraire l'oxygène de l'air, comme la cavité buccale des anguilles électriques. Les gouramis, ainsi que les familles de poissons-chats Channidae, Luciocephalidae et Clariidae ont un « organe labyrinthe » au-dessus des branchies pour cela. Des organes « modifiés », comme les poumons provenant eux-mêmes d'une modification de la vessie natatoire, sont présents chez les Cobitidae, les trahiras et certains poissons-chats des familles Loricariidae, Callichthyidae et Scoloplacidae qui peuvent absorber l'air à travers leur système digestif. Certaines espèces, tels les dipneustes, possèdent des poumons plus ou moins rudimentaires qui leur permettent de survivre en respirant hors de l'eau dans des mares appauvries en oxygène ou régulièrement asséchées. Les semionotiformes et le poisson-castor ont une vessie natatoire vascularisée utilisée de la même manière. La faculté de respirer de l'air est surtout utile aux poissons habitant des eaux peu profondes où la concentration en oxygène peut baisser à certains moments de l'année. Pendant ces périodes, les poissons qui dépendent uniquement de l'oxygène contenu dans l'eau (comme les perches) suffoquent rapidement tandis que les poissons pouvant respirer de l'air survivent mieux, même dans une eau qui ressemble davantage à de la boue humide.
Chez les poissons, la nourriture est ingérée par la bouche et subit un début de trituration dans l'œsophage. Le broyage des aliments se fait essentiellement dans l'estomac et, chez de nombreuses espèces, dans des diverticules en forme de doigt appelés cæca pyloriques. Des organes comme le foie et le pancréas apportent de nouvelles enzymes digestives au fur et à mesure que la nourriture progresse dans le tube digestif. Les intestins complètent le processus de digestion et d'absorption des nutriments.
Systèmes Sensoriels et Régulation Thermique
La plupart des poissons possèdent des organes sensoriels très développés. La majorité des poissons diurnes ont des yeux capables d'une vision des couleurs au moins aussi bonne que celle des humains. De nombreux poissons possèdent aussi des cellules spécialisées appelées chémorécepteurs responsables d'un extraordinaire sens du goût et de l'odorat. Le bulbe olfactif, dans la partie la plus antérieure du cerveau, est impliqué dans le sens de l'odorat. Le cerveau stricto sensu, contrairement à celui des autres vertébrés, s'occupe plus du traitement des informations concernant le sens de l'odorat que de la réalisation des mouvements volontaires. Les lobes optiques traitent les informations provenant des yeux. Les poissons présentent des masses globuleuses qui sont des centres olfactifs et visuels. Leur bulbe rachidien est développé, en partie en relation avec l'organe spécial de la ligne latérale.
Bien qu'ils aient des oreilles internes, de nombreux poissons entendent mal les sons. Cependant, la plupart des poissons possèdent des récepteurs sensitifs formant la ligne latérale, qui leur permet de détecter les courants faibles et les vibrations (par exemple le mouvement de leur proie). Les poissons-chats et les requins possèdent de plus des organes qui détectent des courants électriques de faible intensité. Certaines espèces - telles les mormyridés, notamment Gnathonemus petersii - peuvent être utilisées pour détecter la pollution de l'eau grâce à la variation de leurs émissions électriques. Le cervelet coordonne les mouvements du corps, et le bulbe rachidien contrôle le fonctionnement des organes internes.
Des chercheurs de l'université d'Édimbourg (Écosse) travaillant sur la truite arc-en-ciel ont découvert en 2003 que les poissons pouvaient ressentir la douleur. Cette étude a été critiquée par James D. Rose la même année, au motif qu'il manque aux poissons un néocortex pour ressentir la douleur.
La plupart des poissons sont des animaux dits à sang froid (poïkilothermes). Cependant, des études récentes montrent que certaines espèces prédatrices sont capables de maintenir leur température interne au-dessus de celle de l'eau. Cette aptitude a été démontrée chez certains téléostéens - Scombroïdés (marlin, espadon, voilier), gastéro (Gasterochisma melampus) - et chondrostéens - Lamnidés (requin mako, requin blanc, requins taupes). Leur capacité à réguler leur température varie de l'espadon, qui ne réchauffe que ses yeux et son cerveau, au thon bleu et aux requins taupes capables d'élever leur température corporelle de 20 °C par rapport à celle de l'eau. Des retes mirabiles constituent une sorte d'échangeur thermique et permettent de récupérer les calories du sang chaud et pauvre en oxygène. Le sang qui arrive aux organes est donc relativement chaud tout en diminuant le coût énergétique.
Reproduction et Développement chez les Poissons
La reproduction chez les poissons présente une grande diversité. Chez les poissons ovipares, après une fécondation généralement externe, la femelle pond ses œufs, et les embryons se développent et éclosent en dehors de son corps. Les embryons sont nourris grâce aux réserves contenues dans l'œuf. Plus de 97 % des espèces connues de poissons sont ovipares, dont par exemple les saumons, les poissons rouges, les thons, les anguilles, etc. L'abandon des œufs peut se faire au hasard, par dépôt des œufs dans un nid construit ou un abri naturel, par suspension des œufs à un support, par dépôt puis mise à l'abri des œufs dans la bouche d'un des parents, ou par enfouissage des œufs dans les sédiments.
La fécondation est le plus souvent externe, les gamètes se mélangeant à proximité des deux poissons. Chez quelques poissons, la fécondation est interne, le mâle utilisant un organe intromittant pour déposer le sperme dans le cloaque de la femelle. Il s'agit surtout des requins ovipares comme le requin dormeur cornu et des raies ovipares (comme Rajidae). Les jeunes poissons sortant de l'œuf sont appelés larves. Elles sont généralement peu formées, transportent une vésicule vitelline (ou sac vitellin) contenant leurs réserves nutritives et ont une apparence très différente des adultes. La période larvaire des poissons ovipares est généralement courte (quelques semaines), les larves grandissant rapidement tandis que leur structure et leur apparence change (la métamorphose) pour ressembler aux jeunes de leur espèce. On peut distinguer, sur une larve de saumon, les artères autour de la vésicule vitelline, des gouttes d'huile, le système digestif, la principale artère caudale, la vessie et les arcs des branchies.
Chez les poissons ovovivipares, les œufs restent dans le corps de la mère après fécondation interne. Chaque embryon se développe dans son œuf, sans utiliser de réserves nutritives de la mère, puis sort du corps de sa mère lors de l'éclosion. Chez les poissons vivipares, la fécondation est aussi interne mais chaque embryon reçoit ses nutriments du corps de sa mère, et non des réserves de l'œuf. Les petits sont mis au monde par accouchement. Les poissons vivipares ont une structure similaire au placenta des mammifères, reliant le sang de la mère et celui de l'embryon. Parmi les poissons vivipares, on peut noter les Embiotocidae, les Goodeidae et les requins citrons. Les embryons de certains poissons vivipares ont un comportement appelé oophagie, lorsque l'embryon en développement mange les œufs produits par la mère. Ce comportement a été observé chez certains requins comme le requin mako ou le maraîche mais aussi chez d'autres poissons comme le Nomorhamphus ebrardtii. Le cannibalisme in utero est un comportement encore plus inhabituel, lorsque l'embryon le plus grand mange ses frères plus petits et plus faibles.
Le Système Immunitaire des Poissons
Le système immunitaire des poissons varie considérablement selon les groupes taxonomiques, reflétant des millions d'années d'évolution.
Chez les Agnathes (myxines et lamproies), il n'existe pas de véritables organes immunitaires. En revanche, ces poissons possèdent des tissus lymphoïdes, capables de produire des cellules immunitaires, dans leurs organes. Par exemple, les érythrocytes, macrophages et plasmocytes sont produits dans le rein antérieur (ou pronéphros), et certaines zones de l'intestin (où les granulocytes font leur maturation) chez la myxine ressemblent à une moelle osseuse primitive.
Les poissons cartilagineux (requins et raies) ont un système immunitaire plus évolué que les Agnathes. Ils possèdent trois organes spécialisés qui sont typiques des Chondrichtyens : les organes épigonaux, entourant les gonades, qui ressemblent à la moelle osseuse des mammifères ; l'organe de Leydig, à l'intérieur des parois de l'œsophage ; une valve spirale dans l'intestin. Tous ces organes contiennent des cellules immunitaires typiques (granulocytes, lymphocytes et plasmocytes). Les Chondrichtyens possèdent aussi un thymus identifiable et une rate bien développée (qui est leur organe immunitaire le plus important) où divers lymphocytes, plasmocytes et macrophages se développent et sont stockés.
Les Chondrostéens (esturgeons et Polyodontidae) et les polyptères possèdent un site principal de production de granulocytes dans une masse cellulaire associée aux méninges (membranes entourant le système nerveux central), et leur cœur est souvent couvert d'un tissu qui contient des lymphocytes, des fibroblastes et quelques macrophages. Le rein des Chondrostéens est un organe hématopoïétique important où les érythrocytes, granulocytes, lymphocytes et macrophages se développent.
Les Téléostéens, ou poissons osseux ont, comme les Chondrostéens, leurs principaux tissus immunitaires dans les reins (surtout dans le rein antérieur, où l'on trouve de nombreuses cellules immunitaires de type varié). Les Téléostéens possèdent de plus un thymus, une rate et des zones immunitaires éparses intégrées dans des tissus muqueux (par exemple dans la peau, les branchies, l'intestin et les gonades). On pense que comme chez les mammifères, leurs érythrocytes et granulocytes (entre autres les neutrophiles) résident dans la rate, alors que les lymphocytes sont majoritaires dans le thymus. D'après une découverte récente non encore confirmée, un système lymphatique similaire à celui des mammifères a été décrit chez un Téléostéen, le poisson zèbre (Brachydanio rerio). Ce système serait l'endroit où les lymphocytes T s'accumuleraient en attendant d'être mis en contact avec un antigène.
Système Excréteur
Le système excréteur des poissons joue un rôle crucial dans l'élimination des déchets métaboliques et la régulation de l'équilibre hydrique. Certains déchets métaboliques peuvent diffuser directement à travers les branchies, du sang vers l'eau environnante. Les autres sont éliminés par les reins, organes excréteurs qui filtrent les déchets pour les éliminer du sang.
Les poissons d'eau de mer ont tendance à perdre de l'eau à cause des phénomènes d'osmose et les reins concentrent les déchets afin de réabsorber l'eau au maximum. Il se passe l'inverse chez les poissons d'eau douce, qui ont tendance à absorber de l'eau par osmose.
tags: #bulbe #arterielle #chez #le #poisson