La moelle allongée, plus communément appelée bulbe rachidien, représente la partie terminale et inférieure du tronc cérébral. Cette structure anatomique cruciale agit comme une jonction essentielle entre l'encéphale et la moelle épinière, jouant un rôle pivot dans le contrôle de fonctions réflexes vitales pour la survie, tant dans la vie de relation que dans la vie de nutrition. C'est également un carrefour majeur pour les voies sensitives et motrices du névraxe.
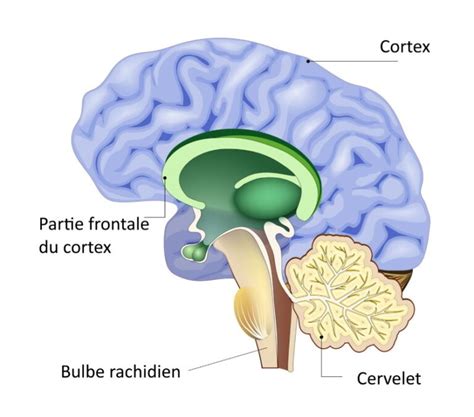
Anatomie Générale du Bulbe Rachidien
Le bulbe rachidien, ou medulla oblongata selon la nomenclature internationale, prolonge sans limites nettes la moelle épinière vers le haut. Il constitue la portion la plus basse, inférieure ou caudale, du tronc cérébral. Embryologiquement, il provient du développement de la vésicule cérébrale la plus postérieure, le myélencéphale. Le bulbe rachidien se situe dans la fosse crânienne postérieure, sous la tente du cervelet. Sa partie rostrale est continue avec le pont supérieur, formant la jonction pontobulbaire.
La surface ventrale, ou antérieure, du bulbe rachidien fait face à la partie basilaire de l'os occipital et au processus odontoïde de l'axis (C2), séparée de ces structures par les méninges et les ligaments des articulations atlanto-occipitale et atlanto-axoïdienne. La surface dorsale, ou postérieure, du bulbe rachidien fait face au quatrième ventricule du cerveau.
Les surfaces ventrale et dorsale du bulbe rachidien sont marquées par des protubérances et des fissures uniques, formées par la présence de différents noyaux et de leurs faisceaux associés.
Face Ventrale
Le long de la ligne médiane de la surface ventrale de la moelle allongée se trouve la fissure médiane antérieure. Il s'agit du prolongement de la fissure médiane antérieure de la moelle spinale. De chaque côté de la fissure se trouve une protubérance verticale connue sous le nom de pyramide médullaire, formée par les fibres du faisceau corticospinal. Chaque moitié de la surface ventrale de la moelle allongée présente deux sillons : le sillon antérolatéral et le sillon postérolatéral. Entre les sillons, et juste latéralement et postérieurement à chaque pyramide, se trouve une autre structure ovale connue sous le nom d'olive. L'olive est formée par la présence du noyau olivaire inférieur dans la moelle allongée. La face ventrale est marquée par un sillon médian masqué, vers le bas, par les fibres entrecroisées de la décussation pyramidale. Il sépare deux cordons blancs longitudinaux, prolongements des cordons antérieurs médullaires : les pyramides antérieures, limitées latéralement par le sillon collatéral antérieur ou préolivaire.
Faces Latérales
Les faces latérales, ou cordons latéraux, prolongent celles de la moelle. À leur partie supérieure, on distingue l'olive bulbaire ou inférieure, un important renflement ovoïde d'à peu près 1 cm de grand axe sur 0,5 cm. La limite dorsale est formée par le sillon collatéral postérieur.
Face Dorsale
La surface dorsale de la moelle allongée est marquée par le sillon médian dorsal sur la ligne médiane. De chaque côté du sillon médian dorsal, on trouve deux protubérances verticales parallèles au sillon. Celle qui est située immédiatement latéralement au sillon est formée par le faisceau gracile qui monte depuis la moelle spinale. La partie crâniale du faisceau gracile contient le noyau gracile, un noyau relais qui établit une synapse avec les fibres du faisceau gracile. Le noyau gracile forme un élargissement visible sur la surface dorsale de la moelle allongée crâniale, appelé tubercule gracile. Latéralement à chaque faisceau gracile se trouve une autre protubérance verticale formée par le faisceau cunéiforme. Latéralement à chaque noyau cunéiforme se trouve une autre protubérance appelée tubercule trigéminal, qui est formée par le noyau spinal du nerf trijumeau (V).
La face postérieure, plus complexe à décrire, comprend en fait deux portions, une inférieure et une supérieure. La portion inférieure continue simplement les cordons postérieurs de la moelle. On peut ainsi y distinguer, de part et d'autre du sillon longitudinal dorsal, la portion bulbaire du faisceau de Goll (fasciculus gracilis n.i.) se renflant à sa partie supérieure en une petite éminence (clava), et, plus latéralement, celle du faisceau de Burdach (fasciculus cuneatus n. i.) se dilatant un peu, vers le haut, en tubercule cunéiforme. Celui-ci se poursuit ensuite sans limites tranchées dans le corps restiforme, ou pédoncule cérébelleux inférieur, qui se rattache au cervelet. À peu près au milieu de la hauteur du bulbe, les lèvres du sillon longitudinal s'écartent et laissent voir, à travers la toile choroïdienne inférieure, une portion du plancher du quatrième ventricule, que l'on décrit classiquement comme la portion supérieure de la face postérieure du bulbe. La décrire revient à décrire le triangle inférieur du plancher du quatrième ventricule, qui, de forme losangique irrégulière, apparaît limité, en ses côtés inférieurs, par les corps restiformes et, en ses côtés supérieurs, par les pédoncules cérébelleux supérieurs.
Organisation Interne du Bulbe Rachidien
Sur une coupe transversale de la moelle allongée, on peut identifier trois parties du ventral au dorsal : la base, le tegmentum et le tectum. La base de la moelle allongée contient la décussation pyramidale du faisceau corticospinal. Le tegmentum de la moelle allongée abrite les noyaux olivaires inférieurs, ainsi que les noyaux des nerfs crâniens IX à XII. Ainsi, divers noyaux (substance grise) se trouvent principalement dans la partie dorsale de la moelle allongée, tandis que les faisceaux (substance blanche) passent principalement par la partie ventrale. Un noyau est un groupe de corps cellulaires neuronaux au sein du système nerveux central. Les noyaux et les faisceaux sont des structures tridimensionnelles qui s'étendent généralement sur plusieurs niveaux.
Les Noyaux du Bulbe Rachidien
Les noyaux de la moelle allongée sont impliqués dans de nombreuses fonctions corporelles importantes. Imaginez trancher la moelle allongée transversalement au niveau du nerf vague (X). Comme dans le reste du tronc cérébral, la majorité des noyaux de la moelle allongée se trouvent dans sa moitié dorsale. Tous les noyaux sont répartis bilatéralement, ce qui signifie qu'il y a un noyau de chaque côté de la ligne médiane.
- Noyaux du raphé : Ces nombreux noyaux appartiennent à la formation réticulée de la moelle allongée.
- Noyau périhypoglosse (de Roller).
- Noyau hypoglosse : Ce noyau est représenté dans le myélencéphale par les neurones du nerf crânien XII (nerf hypoglosse) qui innervent les 4 myotomes occipitaux et la musculature de la langue.
- Noyau dorsal du nerf vague : Ce noyau, qui fait partie de la colonne efférente viscérale générale (EVG), contient les neurones préganglionnaires du système parasympathique et innerve la musculature cardiaque et viscérale involontaire.
- Noyau vestibulaire médial (de Schwalbe) : Ce noyau fait partie du système vestibulaire et s'étend du pont caudal à la moelle allongée rostrale. Il reçoit des informations des canaux semi-circulaires de l'oreille interne, puis les projette via des fibres qui rejoignent le faisceau longitudinal médian (FLM).
- Noyau cunéiforme : Ce noyau reçoit des informations sensitives concernant le toucher léger, la proprioception et les vibrations du membre supérieur ipsilatéral, transportées par le faisceau cunéiforme. Dans la partie caudale « fermée » du myélencéphale, les neuroblastes des plaques alaires dorsales migrent dans la zone marginale pour former les noyaux gracile (médian) et cunéiforme (latéral), relais de la sensibilité proprioceptive et épicritique vers le cervelet et le thalamus.
- Noyau spinal du trijumeau : Ce noyau est également représenté par une protubérance appelée tubercule trigéminal.
- Noyau ambigu : Ce noyau fait partie du groupe respiratoire ventral.
- Noyau réticulaire latéral : Ce noyau reçoit et intègre les influx provenant de diverses sources, qu'il transmet ensuite à l'hémisphère ipsilatéral du cervelet.
- Noyaux olivaires : Ces noyaux comprennent le noyau olivaire accessoire dorsal, le noyau olivaire inférieur et le noyau olivaire accessoire médial. Le noyau de l'olive bulbaire est constitué par des neurones quittant la lame alaire pour migrer dans la partie ventrale du bulbe. C'est la première structure supra-segmentaire à se mettre en place.
Les lames fondamentales du rhombencéphale constituent dès le 28e jour les noyaux moteurs des nerfs crâniens V à XII. Vers la 5e semaine, les lames alaires constitueront les noyaux sensitifs correspondants. Ces noyaux sont répartis en sept groupes, les groupes végétatifs étant situés en regard du sulcus limitans et les groupes somatiques sensitifs latéralement et moteurs médialement. L'amincissement extrême de la paroi du tube neural dans sa région dorsale conduit à l'accolement de l'épithélium épendymaire (Lamina epithelialis) et des leptoméninges (pie-mère + arachnoïde) pour former la toile choroïdienne au niveau de laquelle se différencient les plexus choroïdes. À l'opposé du toit, le plancher et les parois latérales s'épaississent et on retrouve à ce niveau les structures des lames fondamentale et alaire qui sont mises « à plat » séparées par le sulcus limitans. Ce phénomène a pour conséquence l'isolement sous forme de noyaux (dont certains très longs) des lames de substance grise qui formaient une unité structurelle dans la moelle.
Les Faisceaux du Bulbe Rachidien
- Faisceau pyramidal : C'est le faisceau le plus ventral de la moelle allongée. C'est le faisceau même qui se projette dans les pyramides visibles sur la surface ventrale de la moelle. La voie pyramidale, ou voie corticospinale, est impliquée dans la motricité volontaire.
- Lemnisque médian : Il passe juste dorsalement au faisceau pyramidal. Il établit une synapse avec les fibres provenant des noyaux cunéiforme et gracile qui transmettent les informations somatosensorielles du corps.
- Pédoncule cérébelleux inférieur : Il passe latéralement au faisceau longitudinal médian. Il s'agit d'une masse de substance blanche qui contient les faisceaux reliant la moelle spinale au tronc cérébral et au cervelet.
- Tractus spinal du nerf trijumeau : Il passe ventralement au pédoncule cérébelleux inférieur.
- Tractus spinocérébelleux : Il se trouve en avant du noyau lenticulaire latéral.
- Tractus spinothalamique : Il passe médialement et ventralement au tractus spinocérébelleux. Il transmet les informations relatives à la douleur et à la température du côté controlatéral du corps au thalamus.
La Décussation Pyramidale : Un Phénomène Crucial de la Motricité Volontaire
La décussation est l'entrecroisement des fibres nerveuses symétriques droites et gauches au niveau de la face dorsale du bulbe rachidien et des pédoncules cérébraux. Au point de croisement, les faisceaux nerveux de la droite passent à gauche et inversement. La décussation, par son entrecroisement de fibres nerveuses sous la forme d'un X, fait que des parties du corps sont contrôlées par la moitié opposée du cerveau.
La décussation des pyramides est un exemple emblématique de ce phénomène. La motricité volontaire est très rapide. Elle est constituée par un neurone qui part du cortex. Les fibres vont se concentrer pour former un faisceau qui va se regrouper au niveau du centre ovale, puis passer par la capsule interne et descendre par le tronc cérébral. Ces éléments vont constituer le faisceau cortico-nucléaire (faisceau géniculé). La caractéristique de ce faisceau est que toutes les fibres croisent la ligne médiane, c'est un faisceau qui est moteur pur. Le deuxième contingent est le faisceau cortico-spinal, dont les fibres naissent également au niveau du cortex moteur (circonvolution frontale ascendante), se concentrent au niveau du centre ovale, passent par la capsule interne et traversent les éléments du tronc cérébral : le mésencéphale, la protubérance et le bulbe. Toutes ces fibres gagnent la moelle épinière (d'où le nom spinal), mais la grande majorité (environ 80%) va, à la partie inférieure du bulbe, croiser la ligne médiane pour constituer le faisceau pyramidal croisé. La voie pyramidale est celle de la motricité volontaire. Elle décusse, donc un hémisphère contrôle l'autre côté.
Le faisceau pyramidal se constitue à partir de grosses cellules : les cellules de Betz, dans un endroit précis : la zone située immédiatement en arrière de la scissure centrale, c'est la circonvolution frontale ascendante limitée en arrière par la scissure de Rolando et en avant par le sillon précentral. Cette zone s'initialise au-dessus de la scissure latérale, parcourt toute la face latérale, les parties supérieure et médiale du cerveau et s'arrête au niveau du sillon frontocalleux. Les fibres vont décrire une couronne rayonnante et vont se rassembler au centre ovale. Là s'opère un mouvement de rotation, les fibres les plus latérales se disposent en dedans, en avant puis elles gagnent la capsule interne (entre, en dedans le thalamus, et en dehors la partie médiane du noyau lenticulaire (le pallidum)). Celles qui vont au genou (F3) se disposent en dedans et en avant et occupent le genou. Tout le faisceau cortico-nucléaire occupe le genou de la capsule interne d'où le nom "faisceau géniculé".
Les fibres du faisceau pyramidal restent antérieures. Là, 80% des fibres du faisceau corticospinal décussent à la partie basse du bulbe pour constituer le faisceau pyramidal croisé. Le faisceau cortico-nucléaire va s'épuiser à la partie basse du bulbe. La racine antérieure est motrice pure. Le faisceau pyramidal direct se dispose en avant en regard du sillon médial antérieur. Le faisceau pyramidal croisé se dispose en avant de la corne postérieure de la moelle. À chaque niveau métamérique, le faisceau pyramidal croisé abandonne des fibres qui vont gagner la corne antérieure de la moelle et là, font relai avec le deuxième neurone, le deutoneurone, qui va amener à la constitution de la racine antérieure de la moelle, racine qui se termine directement sur le muscle. Le faisceau pyramidal direct va, à chaque niveau métamérique, abandonner des fibres qui vont décusser, gagner la tête de la corne antérieure, et là faire relai avec le neurone effecteur (le motoneurone, aussi appelé deutoneurone) qui sortira par la racine antérieure. Le faisceau pyramidal qui n'a pas croisé la ligne médiane au niveau du bulbe le fera dans la moelle épinière.
Neuroanatomie 12-2: Anatomie clinique du système pyramidal (motricité volontaire) [Pyramidal tracts]
Fonctions du Bulbe Rachidien
Le bulbe rachidien abrite des centres nerveux importants qui régulent des fonctions vitales.
Contrôle Respiratoire
Le centre respiratoire est un groupe complexe de noyaux situés dans le pont et la moelle allongée. Il se compose de trois parties : le groupe respiratoire dorsal, le groupe respiratoire ventral et le centre pneumotaxique.
- Groupe respiratoire dorsal : Ce groupe est responsable de l'inspiration ou de l'inhalation de l'air, et il joue le rôle le plus fondamental dans le processus respiratoire. La majorité de ses neurones se trouvent dans le noyau du tractus solitaire, qui reçoit des informations des chémorécepteurs périphériques sur la saturation en oxygène du sang. Après avoir reçu ces informations, le groupe respiratoire dorsal stimule le nerf phrénique pour contracter le diaphragme, ainsi que les nerfs spinaux thoraciques pour contracter les muscles intercostaux.
- Groupe respiratoire ventral : Il est constitué de la partie rostrale du noyau ambigu et d'un petit noyau satellite appelé noyau rétroambigu, qui se trouve en dessous du premier. Ces neurones sont inactifs pendant la respiration normale, non forcée.
- Centre pneumotaxique pontin : Ce centre se trouve dans le noyau parabrachial du pont rostral et est connecté au groupe de neurones respiratoires dorsaux. La fonction principale du centre pneumotaxique est de « désactiver » le signal inspiratoire provenant du groupe respiratoire dorsal, dictant ainsi le cycle respiratoire et la durée de l'inspiration.
Régulation de la Pression Artérielle
Le système de contrôle de la pression artérielle se trouve dans le centre vasomoteur de la moelle allongée.
- Zone vasoconstrictrice : Elle se trouve dans la partie antérolatérale de la moelle rostrale et se connecte aux neurones de la moelle spinale qui contribuent plus tard aux nerfs sympathiques périphériques qui provoquent une vasoconstriction systémique.
- Zone vasodilatatrice : Elle se trouve dans la partie antérolatérale de la moelle caudale. Elle se connecte à la zone vasoconstrictrice et inhibe sa fonction en cas de besoin, provoquant ainsi une vasodilatation systémique.
- Zone sensorielle : Elle se trouve dans le noyau du tractus solitaire et reçoit les stimuli des barorécepteurs du corps aortique via le nerf glossopharyngien (IX).
Autres Fonctions Réflexes
Le bulbe rachidien abrite également des centres gérant des activités telles que la déglutition, la toux, le vomissement, etc. Le réflexe permettant au contenu buccal de passer dans l'œsophage fait intervenir des fibres afférentes provenant des rameaux sensitifs du nerf pneumogastrique et du nerf trijumeau ; elles excitent des centres nerveux bulbaires. De nombreuses drogues perturbent l'activité des noyaux bulbo-mésencéphaliques impliqués dans la régulation de la vie neurovégétative.
Vascularisation du Bulbe Rachidien
La surface de la moelle allongée est irriguée par les branches des artères vertébrales et basilaires : les artères spinales antérieure et postérieure, et les artères cérébelleuses postéro-inférieures.
Pathologies Associées au Bulbe Rachidien
Les atteintes vasculaires de la moelle allongée peuvent donner lieu à une condition connue sous le nom de syndrome médullaire latéral de Wallenberg. Il est causé par une obstruction de l'artère vertébrale ou de l'artère cérébelleuse postéro-inférieure (PICA), qui irriguent la partie dorsolatérale de la moelle. Une atteinte vasculaire de l'artère spinale antérieure ou de l'artère vertébrale peut également affecter le flux sanguin à travers les artères médullaires directes qui irriguent la moelle médiale.
Un autre phénomène lié à l'interruption des stimulations nerveuses est la sensation de picotement intense. Lorsque les fibres nerveuses motrices et sensitives sont pincées, les commandes motrices ne peuvent passer et le cerveau ne reçoit plus les stimulations sensitives. Lorsque la pression est relâchée, les stimulations nerveuses reprennent toutes en même temps, provoquant une douleur, d'où la sensation de picotement intense qui s'ensuit.
Développement Embryonnaire du Myélencéphale
Le myélencéphale est la subdivision la plus caudale du rhombencéphale. Il va former le bulbe rachidien (Medulla oblongata) chez l'adulte. Le myélencéphale abrite la plupart des noyaux des nerfs crâniens ainsi que les centres qui contrôlent la respiration, le rythme cardiaque et divers centres gérant des activités tels que la déglutition, la toux, le vomissement, etc.
Dans la partie caudale « fermée » du myélencéphale, les neuroblastes des plaques alaires dorsales migrent dans la zone marginale pour former les noyaux gracile (médian) et cunéiforme (latéral), relais de la sensibilité proprioceptive et épicritique vers le cervelet et le thalamus. Quant à la partie ventrale du myélencéphale, elle est le lieu de passage dès le 4e mois de la voie pyramidale (ou voie corticospinale impliquée dans la motricité volontaire).
Dans la partie rostrale « ouverte » du myélencéphale, la courbure pontique (Formation des vésicules cérébrales secondaires et des courbures) de concavité dorsale va dilater le toit de la cavité épendymaire sous-jacente qui forme alors un losange. Le toit, du fait de cet étirement latéral, devient très mince constituant la toile choroïdienne du 4e ventricule. Il s'agit d'une couche de cellules épendymaires cubiques tapissée par le mésenchyme très vascularisé de la pie-mère. La toile choroïdienne va proliférer sur les bords latéraux du ventricule pour former les plexus choroïdes qui ont pour fonction la sécrétion du liquide céphalo-rachidien (LCR) à partir du sang. Les parois latérales commencent à s'écarter, avec pour conséquence l'étirement et l'amincissement de la plaque du toit. La plaque du toit s'est étirée et recouvre la cavité du 4e ventricule. Conséquence de l'écartement des parois, les lames alaires et fondamentales se trouvent quasiment à l'horizontale, le sillon limitant séparant toujours ces deux entités.
L'amincissement extrême de la paroi du tube neural dans sa région dorsale conduit à l'accolement de l'épithélium épendymaire (Lamina epithelialis) et des leptoméninges (pie-mère + arachnoïde) pour former la toile choroïdienne au niveau de laquelle se différencient les plexus choroïdes. À l'opposé du toit, le plancher et les parois latérales s'épaississent et on retrouve à ce niveau les structures des lames fondamentale et alaire qui sont mises « à plat » séparées par le sulcus limitans. Ce phénomène a pour conséquence l'isolement sous forme de noyaux (dont certains très longs) des lames de substance grise qui formaient une unité structurelle dans la moelle. L'étirement de la plaque du toit et sa diminution d'épaisseur a formé la toile choroïdienne (Velum medullare) dans lequel se formera de chaque côté un plexus choroïde. À ce stade sept paires de noyaux moteurs (médians), viscéraux (paramédians, sous le sulcus limitans) et sensitifs (latéraux) s'organiseront dans les lames alaires et fondamentales.
Le Cervelet et ses Connexions avec le Bulbe Rachidien
La structure du cervelet (ou "petit cerveau") est similaire à celle du télencéphale. Les neurones, appelés neurones pyramidaux (certains ont un diamètre approchant 0,1 mm !), vont exciter nos muscles et produire le mouvement. Ils sont très importants pour la motricité car c'est eux qui génèrent l'ordre pour le mouvement volontaire. Le cortex moteur, situé dans le lobe frontal du cerveau, est la zone de départ de la voie pyramidale. On peut encore mieux apprécier ses limites latérales sur une vue en coupe horizontale qui permet de visualiser les deux bras qui la composent entre thalamus (th), noyau lenticulaire (nl) et noyau caudé (nc).
Le cervelet est connecté au tronc cérébral par des faisceaux de fibres nerveuses qui entrent dans le cervelet. Il s'agit d'une succession de petits segments. Ces faisceaux sont importants pour la conduction nerveuse. Les cellules de Purkinje, un type de neurone du cervelet, sont des neurones très ramifiés qui reçoivent cet influx nerveux le long de leur axone vers les noyaux cérébelleux. Les noyaux cérébelleux sont des groupes de corps cellulaires neuronaux au sein du cervelet. La direction des fissures qui séparent les lobules cérébelleux est grossièrement transversale.
Le pédoncule cérébelleux inférieur, qui contient les faisceaux reliant la moelle spinale au tronc cérébral et au cervelet, assure une communication essentielle entre ces structures. Les informations sensitives, y compris la proprioception et le toucher, ascendantes depuis la moelle épinière passent par le bulbe rachidien pour atteindre le cervelet, notamment via les noyaux gracile et cunéiforme.
tags: #bulbe #rachidien #decussation #yramidal