L'étude de l'hérédité des caractères chez les plantes, et plus particulièrement des couleurs florales, révèle la complexité fascinante des mécanismes génétiques. L'Hibiscus, avec sa diversité de teintes, offre un terrain propice à l'exploration des principes de la génétique, des lois fondamentales de Mendel aux phénomènes plus nuancés comme la codominance et l'épistasie.

Les fondements de la génétique : L'héritage de Gregor Mendel
Gregor Mendel, moine catholique, botaniste et généticien, est universellement reconnu comme le père fondateur de la génétique. Ses travaux, menés au XIXe siècle sur des plants de petits pois, ont révolutionné la compréhension de l'hérédité, qui était alors communément admise comme un simple mélange des caractères parentaux.
La démarche expérimentale de Mendel
Mendel a centré son travail sur le Pois (Pisum sativum), un organisme judicieusement choisi pour plusieurs raisons : il présente un grand éventail de couples de caractères alternatifs bien définis (couleur et forme des fruits et des graines mûres, couleur des pétales, taille de la plante, etc.), la structure de sa fleur permet l'autofécondation mais aussi la fécondation croisée contrôlée, son temps de génération est court et les descendants sont nombreux, et les hybrides sont aussi fertiles que les parents.
Sa démarche expérimentale rigoureuse reposait sur cinq points clés :
- Utilisation de lignées pures : Mendel a multiplié ses plantes par autofécondation pendant deux ans pour s'assurer que les descendants présentaient toujours les mêmes caractères.
- Croisements contrôlés : Le croisement de départ (génération parentale) impliquait des plantes qui ne différaient que par un (monohybridisme) ou deux (dihybridisme) caractères.
- Croisements réciproques et test-cross : Il a toujours réalisé des croisements réciproques et des croisements test.
- Analyse de la descendance sur plusieurs générations : Il a étudié la descendance par autofécondation de chaque hybride sur plusieurs générations.
- Généralisation des résultats : Il a réalisé les mêmes séries d'expériences avec sept paires de caractères alternatifs.
Les "particules" de Mendel et les lois de l'hérédité
Mendel a constaté qu'en génération hybride F1 (issue de la fécondation de gamètes de parents P1 et P2), aucun individu ne présentait de mélange de caractères. Par exemple, en croisant des plants de petits pois à graines lisses (P1) avec des plants à graines ridées (P2), tous les individus F1 présentaient des graines lisses. Ce caractère "pois lisse" était donc dominant sur le caractère "pois ridé".
La réapparition du caractère "pois ridé" en génération F2 (issue du croisement de deux individus F1) a permis à Mendel de mettre en évidence que l'hérédité était transmise par des "particules", aujourd'hui appelées allèles. Chaque plant, issu de deux parents, possède ces particules en double exemplaire. Les deux particules se séparent (ségrègent) lors de la formation des gamètes, de sorte que chaque gamète ne possède qu'une seule particule sur les deux.
Résumé des trois lois de Mendel
C'est ainsi qu'il a formulé ses lois fondamentales :
- Première loi de Mendel (Loi de la ségrégation des allèles) : Les deux allèles d'un gène déterminant un caractère se disjoignent (ségrègent) lors de la formation des gamètes : une moitié des gamètes contient l'un des allèles et l'autre moitié contient l'autre. Cette loi résulte du fait qu'il y a ségrégation des chromosomes homologues lors de l'anaphase de la première division méiotique (anaphase I).
- Seconde loi de Mendel (Loi de la dominance) : Si les descendants d'un croisement impliquant deux lignées pures différant par un seul caractère présentent tous le même phénotype, ce phénotype (caractère) est qualifié de dominant (l'autre caractère est dit récessif).
Mendel a également étudié la transmission de deux caractères à la fois. Par exemple, en croisant des plants de petits pois à graines jaunes et lisses (souches pures, génotype AABB) avec des plants à graines vertes et ridées (souches pures, génotype aabb), il a constaté que les particules associées aux deux caractères n'étaient pas forcément transmises ensemble. Elles pouvaient se disjoindre pour s'associer différemment, faisant apparaître de nouvelles combinaisons de caractères ou phénotypes en F2. Ceci a mené à la troisième loi, bien que non explicitement formulée par Mendel comme telle :
- Troisième loi de Mendel (Loi de l'assortiment indépendant des caractères) : Les allèles de gènes différents se répartissent indépendamment les uns des autres lors de la formation des gamètes, à condition que ces gènes soient situés sur des chromosomes différents (gènes indépendants).
Au-delà de la dominance et de la récessivité : La génétique des couleurs florales de l'Hibiscus
Si les lois de Mendel décrivent des principes fondamentaux, l'étude des couleurs florales, notamment chez l'Hibiscus, révèle des interactions alléliques et géniques plus complexes que la simple dominance complète.
Dominance incomplète et codominance
Mendel avait mis en évidence la dominance et la récessivité des allèles en étudiant les couleurs des fleurs de petits pois : un croisement entre un plant aux fleurs roses (P1) et un plant aux fleurs blanches (P2) donnait systématiquement des fleurs roses en F1, indiquant que l'allèle B responsable du caractère rose est dominant sur l'allèle b responsable du caractère blanc.
Cependant, d'autres plantes, comme certaines variétés de roses ou les Ketmies de Syrie (Hibiscus syriacus), montrent des phénomènes différents. Le croisement d'une rose rouge et d'une rose blanche peut donner une couleur rose en F1. De même, un croisement entre des Hibiscus aux fleurs rouges et aux fleurs blanches peut donner des fleurs roses en F1. Dans ces cas, la couleur en F1 n'existe ni chez le parent 1 ni chez le parent 2 et semble provenir d'un mélange des deux. Si l'on croise deux de ces fleurs roses F1, à la seconde génération "F2", on obtiendra des plantes à fleurs rouges, d'autres à fleurs blanches et d'autres à fleurs roses, soit sensiblement une proportion 1/4 (rouge) - 2/4 (rose) - 1/4 (blanc).
Dans cet exemple, les allèles blanc et rouge ont une importance équivalente dans la détermination du phénotype floral : on dit qu'ils sont codominants ou isodominants. C'est un cas de dominance incomplète ou semi-dominance, où l'hétérozygote présente un phénotype intermédiaire entre ceux des deux homozygotes correspondants. L'Hibiscus syriacus est un exemple typique où la variété 'Blue Bird' (bleue), la variété 'Woodbridge' (rouge) et l'hybride obtenu, de couleur intermédiaire, associent les caractères parentaux.

Gènes liés et gènes indépendants
Les gènes responsables des caractères peuvent être situés sur des chromosomes différents (gènes indépendants) ou sur le même chromosome (gènes liés). Quand deux gènes sont portés par des chromosomes différents, une virgule est placée entre les codes utilisés pour les gènes (ex: (A//B)). Quand les gènes sont portés par le même chromosome, on ne met pas de virgule et les lettres utilisées pour coder les gènes sont écrites l'une à côté de l'autre (ex: (AB)).
Dans le cas de l'étude de deux caractères, si les gènes sont indépendants, il y a 4 possibilités de gamètes différents à l'origine de 9 génotypes de fœtus différents. Par exemple, pour les plants de petits pois à graines jaunes et lisses (AABB) croisés avec des plants à graines vertes et ridées (aabb), les gamètes produits par les individus F1 (AaBb) seront AB, Ab, aB, ab dans des proportions équiprobables.
Épistasie et interactions géniques complexes
La couleur des fleurs peut être déterminée par des interactions plus complexes entre plusieurs gènes, un phénomène appelé épistasie. Dans certains cas, l'expression d'un gène peut masquer ou modifier l'expression d'un autre gène situé à un locus différent.
Mendel lui-même a rencontré ces phénomènes "énigmatiques" lors de ses études sur les haricots à fleurs pourpres (Phaseolus multiflorus) croisés avec des haricots à fleurs blanches (Phaseolus vulgaris). Les hybrides F1 étaient tous à fleurs pourpres, mais en F2, il n'obtenait qu'une seule plante à fleurs blanches et une série de couleurs allant du pourpre au violet pâle, loin de la proportion 3 pourpres : 1 blanc attendue pour un caractère dominant/récessif simple. Il a émis l'hypothèse que la couleur des fleurs du P. multiflorus était composée de plusieurs caractères indépendants (A1 + A2 +…) donnant un effet d'ensemble rouge pourpre. Aujourd'hui, on sait que les pigments floraux, comme les anthocyanes, dérivent de voies de biosynthèse complexes impliquant de nombreux gènes qui produisent différentes molécules responsables des teintes orange, rose, rouge, pourpre, violet, bleu.
Un exemple d'épistasie peut être observé lorsque des gènes différents agissent sur la même voie de biosynthèse des pigments. Par exemple, si la couleur est déterminée par la présence d'au moins un allèle "A" et un allèle "B", un croisement parental AAbb x aaBB donnera 100% de AaBb, tous colorés. En revanche, un croisement entre hybrides AaBb donnera 9/16 d'individus colorés (possédant au moins un "A" et un "B") et 7/16 de blancs.
L'influence de l'environnement et la plasticité phénotypique
Au-delà des gènes eux-mêmes, d'autres facteurs peuvent influencer la couleur des fleurs d'Hibiscus. L'expression des gènes peut être modulée par l'environnement (plasticité phénotypique) ou par la présence d'autres gènes donnant des protéines de régulation différentes.
Par exemple, la couleur d'un pigment floral, comme celui du bleuet ou du coquelicot, peut varier en fonction de l'acidité (pH intracellulaire) de la plante, même si le sol est le même. Ainsi, un hibiscus peut ne pas avoir la même teinte de bleu que celui du voisin, même si leurs patrimoines génétiques sont similaires. Des problèmes tels que l'absence ou la modification d'une enzyme dans une chaîne de biosynthèse des pigments peuvent également altérer la couleur finale.
Applications de la génétique en horticulture : Sélection et hybridation
Les connaissances en génétique sont aujourd'hui utilisées pour orienter les productions de plantes à caractères spécifiques, répondant ainsi à la demande des consommateurs. Pour l'Hibiscus, cela permet de créer de nouvelles variétés avec des couleurs, des formes de corolle (ouverte, par exemple) ou d'autres caractéristiques désirables.
Hybridation et création de nouvelles variétés
L'hybridation, c'est-à-dire le croisement entre différentes variétés ou espèces, est une méthode clé pour introduire de nouveaux caractères ou combiner des caractères existants. En choisissant des parents avec des allèles spécifiques, les sélectionneurs peuvent prédire la probabilité d'obtenir certaines couleurs ou autres traits chez la descendance.
Par exemple, en croisant des plants d'Hibiscus aux fleurs bleues et blanches, la probabilité d'obtenir des teintes intermédiaires (roses ou lavande) ou des retours aux couleurs parentales dépendra des allèles impliqués et de leurs interactions (dominance, codominance). Le hasard joue un rôle important dans la répartition des caractères dans les descendances d'hybrides, mais une compréhension des principes génétiques permet de rationaliser la sélection.
Greffage et modification des couleurs
Le greffage, bien que n'étant pas une méthode de modification génétique de la plante porte-greffe, permet d'associer des parties de plantes de génomes différents. Par exemple, greffer trois branches d'Hibiscus de couleurs différentes (rose, blanc et bleu) sur un même porte-greffe n'aura pas d'impact sur la couleur des fleurs produites par le porte-greffe lui-même, qui continuera à produire des fleurs de sa couleur d'origine. En revanche, chaque greffon produira des fleurs de sa propre couleur, car les greffons conservent leur génome. C'est une technique qui permet d'obtenir un arbuste "multicolore" sans modifier le patrimoine génétique des cellules reproductrices.
Résumé des trois lois de Mendel
Le vocabulaire de la génétique et ses nuances
Pour comprendre pleinement la génétique des couleurs et l'héritage des caractères, il est essentiel de maîtriser le vocabulaire spécifique.
- Allèle : Une forme alternative d'un gène. Par exemple, pour la couleur des fleurs, il peut y avoir un allèle pour le rouge et un allèle pour le blanc.
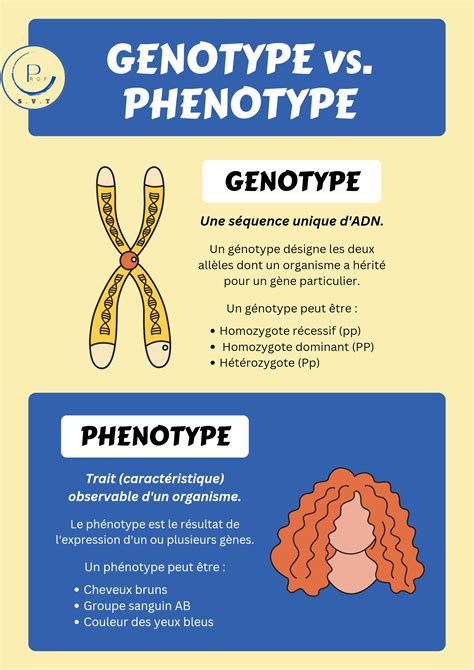
- Génotype : L'ensemble des allèles possédés par un individu pour un ou plusieurs gènes. Il s'écrit toujours entre parenthèses, par exemple (BB) pour homozygote dominant, (Bb) pour hétérozygote, (bb) pour homozygote récessif.
- Phénotype : L'ensemble des caractères observables d'un individu, résultant de l'expression de son génotype et de l'influence de l'environnement. Il s'écrit entre crochets, par exemple [Rouge] pour une fleur rouge.
- Homozygote : Un individu qui possède deux allèles identiques pour un gène donné (par exemple, (BB) ou (bb)).
- Hétérozygote : Un individu qui possède deux allèles différents pour un gène donné (par exemple, (Bb)).
- Dominant : Un allèle qui s'exprime phénotypiquement même lorsqu'il est présent en un seul exemplaire chez un hétérozygote.
- Récessif : Un allèle qui ne s'exprime phénotypiquement que lorsqu'il est présent en deux exemplaires chez un homozygote.
- Codominant (ou isodominant) : Des allèles qui s'expriment tous les deux entièrement chez l'hétérozygote, produisant un phénotype qui combine les caractères des deux allèles. Le groupe sanguin AB est un exemple chez l'homme, où les allèles A et B sont codominants.
- Chromosomes homologues : Une paire de chromosomes, l'un hérité de la mère et l'autre du père, qui contiennent les mêmes gènes aux mêmes loci (emplacements).
- Gamète : Une cellule sexuelle (spermatozoïde ou ovule) qui contient un seul exemplaire de chaque chromosome (haploïde).
- Diploïde : Se dit d'une cellule qui contient deux jeux de chromosomes (un du père et un de la mère).
- Haplotype : Une combinaison d'allèles de gènes liés sur le même chromosome, héritée ensemble.
Exemples d'application de ces concepts
- Fibrose kystique : Cette maladie dépend d'un allèle récessif (k). Un individu atteint a un génotype (kk), tandis qu'un individu sain peut être (KK) ou (Kk). Si deux parents sains sont tous deux porteurs (Kk), ils peuvent avoir un enfant malade avec une probabilité de 1/4. La femme de la première génération, le premier individu en partant de la gauche, a un génotype Kk. L'allèle K est la forme normale du caractère, non malade, et k l'allèle récessif déterminant la maladie. Cette femme est normale, elle possède donc au moins un allèle K. L'homme de la deuxième génération, le quatrième individu en partant de la gauche, est également Kk. Cet homme est normal et possède donc au moins un allèle K. La femme de la troisième génération, le premier individu en partant de la gauche, est Kk. Cette femme est normale et possède donc au moins un allèle K.
- Radis : La forme des radis peut être ronde (RR), longue (LL) ou ovale (RL). C'est un exemple de codominance où les allèles R et L s'expriment tous deux. Si des plantes à radis ronds (RR) sont croisées avec des plantes à radis longs (LL), les descendants F1 seront tous à radis ovales (RL). Si ces F1 sont croisés entre eux, la génération F2 donnera 1/4 de ronds, 1/2 d'ovales et 1/4 de longs. 156 plantes à radis ovales ont été obtenues dans une descendance, confirmant cette proportion.
- Hémophilie et daltonisme : Ce sont des maladies liées au sexe, portées par le chromosome X. Un gène récessif h lié au sexe retarde le temps de coagulation du sang et est responsable de l'hémophilie. Si André est non hémophile, son phénotype est nécessairement XHY, car comme tout homme, il ne possède qu'un allèle H pour ce caractère, porté par la portion du chromosome X non homologue à Y, et qui détermine son phénotype. Bernadette a nécessairement un allèle normal H, puisqu'elle est de phénotype non hémophile, mais aussi un allèle h puisqu'elle l'a donné à Denis, son garçon hémophile. Bénédicte a nécessairement hérité de André un XH, mais elle a une (mal)chance sur deux d'avoir reçu un XH de sa mère Bernadette, et une chance sur deux d'en avoir hérité un Xh. Bénédicte est donc XHXH ou XHXh, de façon équiprobable. Si son premier enfant est hémophile, et comme son mari ne l'est pas, on est certain qu'elle porte un Xh, et qu'elle est donc de génotype XHXh. Elle a donc une (mal)chance sur deux de donner un Xh à son enfant, et une chance sur deux de donner naissance à un garçon. Charlotte a nécessairement hérité de André un XH, mais elle a une (mal)chance sur deux d'avoir reçu un XH de sa mère Bernadette, et une chance sur deux d'en avoir hérité un Xh. Bénédicte est donc XHXH ou XHXh, de façon équiprobable. Elle a donc 3 chances sur 4 de transmettre à son enfant l'allèle H.
- Calvitie : Un caractère qui est dominant chez l'homme et récessif chez la femme.
- Groupe sanguin ABO : L'allèle O est récessif, tandis que les allèles A et B sont dominants sur O et codominants entre eux. Un individu de génotype (A//B) sera du groupe [AB].
- Groupe sanguin MNSs : Les allèles M et N sont codominants au locus de la glycophorine A sur le chromosome 4. Les sous-groupes S et s sont déterminés par deux allèles codant la glycophorine B, où S est dominant sur s. Les deux loci, MN et Ss, sont étroitement liés sur le chromosome n°4, permettant de définir 4 haplotypes possibles : MS, NS, Ms et Ns.
- Hémoglobines : L'allèle (\beta)S, responsable de l'anémie falciforme, peut être considéré comme récessif (pas d'anémie clinique chez les hétérozygotes), dominant (hématies en forme de faucille chez les hétérozygotes dans certaines conditions), ou codominant (présence d'HbA et HbS chez les hétérozygotes), selon le niveau d'analyse du phénotype.
La génétique de la couleur chez l'Hibiscus, bien que difficile à prédire avec une certitude absolue en raison des multiples facteurs en jeu, est une illustration parfaite de la richesse des interactions génétiques, allant des principes fondamentaux de Mendel aux phénomènes complexes de codominance, d'épistasie et d'influence environnementale. La compréhension de ces mécanismes est essentielle pour les horticulteurs et les généticiens qui cherchent à développer de nouvelles variétés aux caractéristiques désirables.
tags: #couleur #hibiscus #croisement #mendel