L'horticulture moderne et l'amélioration des plantes reposent sur une compréhension approfondie de la reproduction végétale, en particulier des mécanismes de l'autofécondation et de l'incompatibilité des croisements. Bien que ces aspects soient souvent délaissés au profit de considérations plus techniques comme la nutrition minérale ou la protection contre les maladies, ils sont fondamentaux pour optimiser la production de fruits et de graines. Cet article vise à apporter des informations rationnelles et incontestables sur ces phénomènes complexes, souvent documentés dans des sources spécialisées et en anglais, afin que le lecteur puisse se forger sa propre opinion.
Les Fondamentaux de la Reproduction Végétale et l'Enjeu de l'Autofécondation
La majorité des espèces d'angiospermes possèdent des organes reproducteurs mâles et femelles sur la même plante, et dans la plupart des cas, au sein de fleurs hermaphrodites. Cette proximité augmente la probabilité d'autofécondation, qui, si elle était généralisée, entraînerait un faible taux de variabilité génétique au sein d'une population. Une telle homogénéité génétique rendrait les plantes plus vulnérables aux variations environnementales, d'où la nécessité de mécanismes évolutifs favorisant les croisements entre individus génétiquement différents.
Historiquement, la sélection variétale a débuté il y a plus de 10 000 ans, lorsque l'homme a commencé à se sédentariser. Il cultivait alors des plantes pour se nourrir et choisissait, après la récolte, les plus belles graines pour les ressemer l'année suivante, inventant ainsi la sélection variétale sans le savoir. À la fin du 19e siècle, les premiers croisements de parents "choisis" ont permis d'identifier les descendants qui correspondaient le mieux aux attentes des agriculteurs, en termes de conservation, de productivité, de goût ou de facilité de culture. Aujourd'hui, l'enjeu est de mettre sur le marché des génétiques avec des caractères intéressants pour répondre à des problématiques nouvelles, ce qui implique une compréhension fine des mécanismes reproductifs des plantes.
L'Auto-Incompatibilité : Un Mécanisme Naturel de Rejet du "Soi"
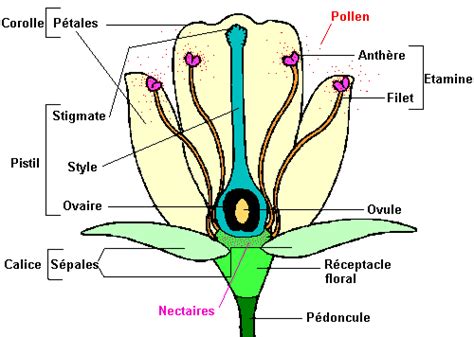
Les mécanismes qui empêchent les autofécondations sont appelés mécanismes d'auto-incompatibilité. Le pollen "soi" ou autopollen est rejeté, tandis que le pollen "non-soi", ou allopollen, est accepté et conduira à une descendance. Ces mécanismes peuvent être très différents. Par exemple, chez certaines primevères, la fleur existe sous deux formes, l'une à styles plus courts que les étamines et inversement. La pollinisation n'aboutit que si les deux partenaires proviennent de formes différentes.
En général, il n'y a aucune différence morphologique entre les fleurs. L'auto-incompatibilité est alors contrôlée par un locus unique nommé S pour "Self-incompatibility". Dans ce contexte, le "soi" et le "non-soi" correspondent respectivement à une identité ou une divergence pour le locus S. Ce locus renferme deux gènes génétiquement liés qui déterminent respectivement la spécificité allélique du pistil ou du pollen. La reconnaissance de l'autopollen est basée sur une interaction entre les produits mâle et femelle du locus S qui aboutit à l'inhibition de croissance du tube pollinique.
Le S-Locus : Support Héréditaire de l'Incompatibilité
Le support héréditaire des mécanismes qui permettent aux plantes de s'entre-croiser ou de se rejeter est simple. Une seule région du génome, le S-locus, porte quelques gènes associés en tandem. Chacun des gènes du tandem a une fonction soit dans le style et stigmate (part femelle), soit dans le pollen (part mâle). Deux de ces gènes, un de chaque partie, encodent chacun une protéine (appelée déterminant), celui de la part femelle et celui de la part mâle. Ils ont une partie variable, plus ou moins longue, et de séquence en bases plus ou moins différentes.
Si les deux parties ont une séquence identique dans la partie variable, alors la réaction d'auto-incompatibilité se déclenchera et le croisement sera incompatible. Ce mécanisme biochimique est particulier pour chaque espèce. Pour l'expérimentateur, tout se passe comme si l'incompatibilité était due à un seul gène, les événements de recombinaison étant exceptionnels dans le S-locus. La diversité entre allèles se crée uniquement par des mutations. Bien que les mutations affectant les S-allèles soient très rares, pour des espèces qui évoluent depuis des millions d'années, on observe une diversité des allèles de l'incompatibilité (S-allèles) dans les populations sauvages.
🧬 Les bases de la génétique
Les Types d'Auto-Incompatibilité : Gamétophytique, Sporophytique et à Action Tardive (LAI)
La diversité se manifeste lors des croisements. Les individus couverts par un sac ne produiront pas de graines (auto-stérile) et très rarement certains pourront en produire (auto-fertile). Lors des croisements des individus deux à deux, une paire est soit compatible soit incompatible. C'est en étudiant de nouvelles paires choisies parmi les individus incompatibles que l'on incrémente le nombre de S-allèles de l'espèce.
Il est possible de distinguer trois types principaux d'auto-incompatibilité, bien que les mécanismes physiologiques exacts de reconnaissance entre le stigmate, le style et le grain de pollen soient souvent inconnus :
Incompatibilité Gamétophytique (GSI) : Quand chacun des grains de pollen d'un individu porte le déterminant protéique de l'allèle de son génome, il est haploïde. Le phénotype du pollen est déterminé par son génotype haploïde (c'est-à-dire par l'allèle qu'il porte). C'est le type le plus fréquent, présent chez la plupart des espèces des Rosaceae et Solanaceae (tabac, tomate sauvage, rose, arbres fruitiers, gueule de loup, coquelicot…). Dans ce système, chaque individu d'une espèce gamétophytique contient toujours deux types de grains de pollen, chacun d'eux portant le déterminant encodé dans son propre génome. En cas d'auto-incompatibilité gamétophytique, la germination du grain de pollen est souvent bloquée sur le stigmate ou la progression du tube pollinique est stoppée dans les tissus femelles après germination.
- Exemple Pédagogique (Urne Gamétophytique) : Imaginez une urne où il y a toujours autant de boules blanches que de boules noires. Avec le système GSI, chaque grain de pollen de l'individu est soit blanc, soit noir, ayant été "peint" après la méiose qui sépare une cellule mère en quatre grains de pollen (deux blancs et deux noirs). Si une variété est inter-incompatible avec une variété X, cela signifie que toutes les variétés inter-incompatibles avec X portent la même paire de S-allèles que X. Toutes les autres combinaisons de paires sont inter-compatibles, car les deux variétés diffèrent entre elles par au moins un des deux S-allèles, et si la réaction d'incompatibilité est déclenchée pour l'allèle en commun, l'autre grain de pollen est compatible. Un grain de pollen sur deux est détruit, l'autre fonctionne.
Incompatibilité Sporophytique (SSI) : Quand chacun des grains de pollen porte les deux déterminants protéiques des deux allèles de l'individu qui l'a produit. Le génome diploïde du parent producteur de pollen détermine le phénotype du grain de pollen. Si la plante émettrice de pollen est hétérozygote pour le locus S, le pollen exprimera deux spécificités alléliques S. Ce type se retrouve chez les Brassicaceae (choux fourrager, brocoli, colza…) et Astéraceae (tournesol…). Chez les Brassicaceae, le rejet se manifeste très tôt, la germination du grain de pollen étant bloquée sur le stigmate.
- Exemple Pédagogique (Urne Sporophytique) : Ici, toutes les boules d'une urne sporophytique sont de la même couleur. Avec le système sporophytique, la cellule mère des grains de pollen a été peinte selon la relation de dominance entre les allèles-S de la plante (sporophyte) qui les produit. Par exemple, comme une carte de donneur de sang, l'olivier a une carte de donneur de pollen. Si Ry est dominant sur Rx, alors le pollen sera couvert du déterminant Ry uniquement. Dans l'olivier, une hiérarchie de dominance existe (ex: R6 > R2 > R1 = R3 = R5 > R4). La dominance s'exprime dans chaque cellule mère des grains de pollen, qui délivre ainsi un seul type de grain de pollen. Par exemple, une variété R1R6 (le pollen est R6) ne peut féconder aucune autre variété qui porte R6 quel que soit l'autre allèle. Cependant, R6 est rare (6 variétés sur 130). Quand le système est sporophytique, comme chaque variété porte deux S-allèles, l'incompatibilité de l'autofécondation ou du croisement est due, respectivement, à un (si dominance) ou deux (si codominance) déterminants. Donc, toutes les paires de variétés qui portent un même S-allèle dominant seront inter-incompatibles, alors que si l'une porte un allèle dominant et l'autre un autre allèle dominant sans autre S-allèle commun, alors elles seront inter-compatibles dans les deux directions. De plus, les paires de variétés qui portent deux S-allèles dominants différents et partagent un même S-allèle récessif donnent des croisements réciproques opposés, l'un incompatible et l'autre compatible.
Incompatibilité à Action Tardive (LAI) : Quand la réaction d'incompatibilité ne se déclenche que très longtemps après la pollinisation. La fécondation se réalise et conduit à un fruit, qui tombera très tardivement, presque à maturité. Cela peut être très frustrant pour le cultivateur, qui ne peut pas relier la chute tardive des fruits à l'incompatibilité du pollen. LAI a été très bien documentée entre 2000 et 2015.
Le Débat autour de la Détermination des Types d'Incompatibilité
Il n'existe pas de tests stricts pour déclarer un système gamétophytique ou sporophytique. Cette difficulté peut conduire à des erreurs d'interprétation. Par exemple, si par malchance, il n'y a pas de paires inter-compatibles entre les individus incompatibles avec une variété X, on pourrait abusivement considérer le système comme gamétophytique alors qu'il est sporophytique. Pourtant, des travaux anciens, comme ceux de Musho (1977) chez Pierre Villemur, ont démontré des croisements complexes : si Picholine et Tanche sont inter-incompatibles, et si Tanche et Bouteillan sont inter-incompatibles, Picholine est un polliniseur de Bouteillan et vice versa.
Les lecteurs des articles scientifiques accessibles par internet peuvent être induits en erreur par des conclusions prématurées tirées de croisements. Il est évident que certains chercheurs ont parfois choisi des paires de variétés pour éviter des différences dans les croisements réciproques, ce qui peut biaiser les conclusions sur le système d'auto-incompatibilité, comme cela a été suggéré pour l'olivier. Nos prédécesseurs chercheurs ont révélé les deux systèmes gamétophytique et sporophytique en étudiant des milliers de croisements dans les années 1940-1950, en observant la germination du pollen sur les stigmates chez plusieurs espèces. Plusieurs dizaines de milliers d'observations ont conduit à l'élaboration de ces deux modèles.
Pour le système des Prunus et des Solanacées, la partie femelle est une S-RNAse dite H. Pour le système sporophytique chez les Brassiceae, il s'agit d'une SRK-kinase dans la partie femelle. L'olivier n'exprime pas de S-RNAse H au niveau de la partie femelle au stade de l'incompatibilité, ce qui a conduit certains chercheurs à en déduire qu'il était de type sporophytique. Paradoxalement, les mêmes chercheurs ont parfois publié simultanément, dans un autre article, que l'olivier était de type gamétophytique, illustrant la complexité et parfois le manque de rigueur dans certaines publications.
L'Auto-Fertilité de l'Olivier et le Modèle des S-Allèles
Chez toutes les espèces qui expriment l'incompatibilité, qu'elle soit de type GSI ou SSI, l'auto-incompatibilité d'un individu est stricte. Or, chez l'olivier, les variétés sont notoirement plus ou moins auto-fertiles. Même si une mutation affectant la partie variable femelle ou mâle d'un S-allèle est très rare, elle peut rendre un individu auto-fertile, ce qui est alors repéré lors d'une expérience d'ensachage par les améliorateurs des plantes. Ils posent des dizaines de milliers de sacs chaque année pour contrôler les croisements. Avant les années 1975, sans ces techniques, l'auto-fertilité des variétés d'olivier n'était pas connue.
Les S-allèles qui ont perdu la faculté d'agir sur l'incompatibilité sont désignés S0. Le modèle des S-allèles a l'avantage de permettre la comparaison de l'auto-fertilité des variétés à la paire d'allèles. Sur plus de 100 variétés élucidées pour leur paire d'allèles, il est apparu que la paire d'allèles est corrélée au niveau d'auto-fertilité. Plus les niveaux de dominance sont éloignés entre les deux S-allèles de la paire, plus la variété est auto-fertile. Ceci est vérifié expérimentalement pour les paires avec R2, R3, R4 et R6.
Cette connaissance est très utile expérimentalement. Si un croisement implique en femelle une paire qui introduit de l'auto-fertilité (comme R4R6, R2R4, ou même R1R5), il faudra vérifier soigneusement que les fruits sont bien dus au croisement par des tests moléculaires de paternité. Quant à l'autofécondation des variétés mâle-stériles détectées avec les marqueurs moléculaires, comme chez Lucques et Olivière, il faut distinguer le résultat de l'interprétation. Un embryon prélevé chez Lucques, puis analysé, peut très bien ne montrer aucun marqueur des variétés que l'on considère comme des polliniseurs possibles.
Tests de Paternité et Vérification des Croisements
Les tests de paternité sont cruciaux pour identifier la variété mâle d'un embryon obtenu sur une variété femelle. La variété hôte, bien identifiée et contrôlée par comparaison du profil moléculaire de l'embryon, permet d'identifier les fragments apportés par un grain de pollen. Un programme informatique analyse les données moléculaires des variétés pour identifier celle qui a donné le profil moléculaire du grain de pollen.
La société SocX, qui réalise ces tests, a construit une base de données des profils moléculaires de près de 200 variétés cultivées en France, incluant quelques variétés à diffusion mondiale. Quand un arbre hôte de la variété d'olivier Roussane, dont le profil moléculaire est connu et dans la base de données de SocX, est utilisé comme parent femelle pour récolter des fruits, puis obtenir des noyaux et enfin des embryons, la comparaison du profil moléculaire de l'embryon et de la mère permet d'identifier les marqueurs présents chez l'embryon et absents chez la mère. Roussane est bien caractérisée par ses 10 paires d'allèles, avec deux tailles d'allèles pour chaque marqueur. Cependant, des controverses peuvent exister, comme la publication récente de Marchese et al. (2015) qui conteste la paire de S-allèles attribuée à deux des variétés (Arbequina et Koroneiki). Si des discordances apparaissent pour les S-allèles portés par des individus de vergers constitués de plants commerciaux, il faudra comparer les profils moléculaires des plants commerciaux à ceux utilisés par des équipes de référence comme Nathalie Moutier, Georgios Koubouris et Daniela Farinelli.
Techniques de Croisement en Horticulture
Pour réaliser un croisement et éviter les autofécondations, il faut tout d'abord castrer les fleurs de la plante qui servira de parent femelle, c'est-à-dire supprimer les étamines. Ensuite, le pollen prélevé sur la plante servant de parent mâle est apporté sur le stigmate. La fleur est ensuite protégée pour éviter des apports de pollen par les pollinisateurs naturels.
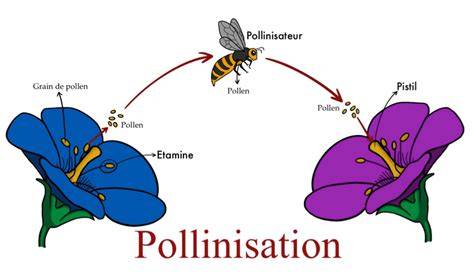
En général, une seule génération de croisement ne suffit pas, surtout lorsque l'un des parents initiaux est très éloigné du type variétal recherché, par exemple une forme sauvage apportant une résistance à une maladie. Il faut alors réaliser plusieurs générations de croisement et de tri, comme l'illustre l'exemple du pommier 'Ariane'.
Ce schéma général est modulé par plusieurs facteurs, en particulier le mode de multiplication et la biologie florale.
Multiplication et Types de Variétés
Plantes à multiplication par graines (la plupart des espèces légumières, pétunias, pensées…) :
- Plusieurs générations d'autofécondation permettent d'obtenir des lignées homogènes.
- Si la variété commerciale est une lignée (haricot, pois, laitue…), l'obtention de quantités importantes de graines de la nouvelle variété est faite en isolement pour éviter d'éventuelles hybridations.
- Si la variété commerciale est un hybride F1 (tomate, piment, concombre, melon, chou-fleur, chicorée witloof, carotte…), il faut produire de grandes quantités de semences avec un système assurant 100% de croisements (ou 0% d'autofécondation). Il peut s'agir de stérilité-mâle (génique ou géno-cytoplasmique), d'auto-incompatibilité (chicorée) ou bien de castration et de pollinisation manuelles. Dans ce dernier cas, le coût de la main d'œuvre est un facteur important du prix des semences, ce qui explique la délocalisation des productions de semences commerciales en Chine ou en Inde pour des espèces comme la tomate ou le piment.
- Si la variété commerciale est une population, l'obtention des semences commerciales est assurée par une multiplication en isolement avec des pollinisateurs naturels.
Plantes à multiplication végétative (arbres fruitiers, vigne, fraisier, pomme de terre, arbustes ornementaux…) :
- On ne réalise pratiquement jamais d'autofécondations contrôlées dans les programmes d'innovation variétale. Dès qu'une plante présente les caractéristiques recherchées, elle peut être multipliée à l'identique par bouturage, greffage, etc., pour être expérimentée à plus large échelle et multipliée commercialement. La nouvelle variété est alors un clone.
- La création de nouvelles variétés de porte-greffe concerne des plantes à multiplication végétative (arbres fruitiers, vigne) mais aussi des espèces à multiplication par graines (tomate, aubergine, melon). Les méthodes sont identiques à l'amélioration des variétés mais les objectifs sont différents : vigueur conférée, résistances aux bio-agresseurs liés au sol, compatibilité entre le porte-greffe et le greffon.
Particularités selon la Biologie Florale
Espèces monoïques (courge, noisetier, noyer…) : Les croisements sont très faciles puisqu'il n'est pas nécessaire de castrer avant d'apporter le pollen du parent mâle, car les fleurs mâles et femelles sont séparées sur la même plante.
Espèces dioïques (asperge, épinard) ou espèces présentant des phénomènes d'auto-incompatibilité (chou, radis) ou de stérilité-mâle : Ces plantes s'autofécondent très difficilement. Il est alors possible de faire des croisements frère x sœur qui permettent également d'atteindre l'homozygotie mais beaucoup plus lentement que les autofécondations successives.
Espèces allogames (oignon, carotte) : Elles présentent un fort effet d'inbreeding, c'est-à-dire une baisse de vigueur et de fertilité au cours des générations successives d'autofécondation nécessaires pour obtenir des lignées fixées.
Variétés triploïdes : Généralement stériles, pour les reproduire par graines il faut faire des hybrides entre un parent diploïde et un parent tétraploïde.
🧬 Les bases de la génétique
Rendement en Graines par Fleur et Outils Biotechnologiques
Le succès d'une pollinisation contrôlée dépend aussi du rendement en graines par fleur :
- Une seule graine chez les Prunus (cerisier, pêcher, abricotier…) ou la laitue.
- Quelques graines chez les Fabacées (pois, haricot), les Brassicacées (chou, radis) ou le pommier et le poirier.
- Plusieurs dizaines à quelques centaines de graines chez les Cucurbitacées (melon, courges) ou les Solanacées (tomate).
Un outil de biotechnologie, l'haploïdie, permet d'atteindre très rapidement l'homozygotie et donc de remplacer plusieurs générations d'autofécondation. Des variétés hybrides F1 de piment, d'aubergine, de melon ont un parent (parfois deux) qui est une lignée haploïde doublée, parfaitement homozygote.
Hybridation Interspécifique
Lorsqu'un caractère recherché n'existe pas dans l'espèce considérée, on peut éventuellement utiliser la variabilité des espèces voisines, sauvages ou cultivées, en réalisant des croisements interspécifiques. La culture in vitro des embryons issus du croisement est souvent nécessaire. Des problèmes de stérilité sont généralement rencontrés dans les premières générations. Pratiquement toutes les résistances aux maladies présentes dans les variétés modernes de tomate proviennent d'espèces sauvages voisines (Lycopersicum peruvianum).
Le Processus de Sélection Variétale et les Biotechnologies
La sélection variétale est un processus de longue haleine. Il commence par le croisement de deux plantes, choisies pour leurs atouts. L'objectif est de stabiliser les caractéristiques identifiées, ce qui est loin d'être facile. Pour réduire ce temps, inhérent à la sélection "classique", les biotechnologies sont des facilitatrices pour réaliser les croisements entre espèces, maîtriser et ajuster les transferts de caractères ou créer plus rapidement des lignées pures. En effet, quand un sélectionneur réalise un croisement entre deux plantes, il brasse un très grand nombre de caractères : ceux qu'il souhaite introduire dans la nouvelle variété, mais également ceux qu'il ne souhaite pas conserver, les "indésirables".
Étapes de la Création Variétale
Croisement des variétés sélectionnées : Différentes techniques permettent de réaliser des croisements artificiels. L'exercice consiste toujours à provoquer la fécondation des ovules d'une plante, choisie pour mère, par le pollen d'une autre, choisie comme père. La démarche varie selon le mode de reproduction des espèces. La synchronisation des cycles de floraison est une étape importante. Le test de compatibilité pollinique entre parents, par la méthode de germination in situ du pollen, permet de vérifier la capacité du pollen à germer sur les stigmates et féconder l'ovule.
Créer, fixer et évaluer les nouvelles plantes : Les graines récoltées à chaque génération sont sélectionnées puis ressemées. À partir de la cinquième génération (F5), les caractéristiques des plantes deviennent plus stables, c'est-à-dire qu'on les retrouve au fur et à mesure des croisements : le brassage des gènes est moins important. Pour la création d'hybrides, l'enjeu est de choisir les parents qui se combinent le mieux. Ces lignées pures et stables servent alors à créer des variétés hybrides, homogènes et reproductibles.
Essais au champ : Après la phase en laboratoire ou serre, les essais se poursuivent au champ, dans différentes régions, pour observer le comportement de chaque variété dans un contexte pédoclimatique donné.
Inscription de la variété : La variété est déposée au CTPS (Comité technique permanent de sélection) et passe deux ou trois années de tests. Seront jugés ses valeurs agronomique, technologique et environnementale (VATE) et ses critères de distinction, d'homogénéité et de stabilité (DSH). Une fois cet examen réussi, la variété est alors multipliée pour être commercialisée sous forme de semences certifiées, un gage de qualité.
Avantages et Inconvénients de l'Hybridation
L'hybridation, pratiquée depuis des millénaires, est au cœur du travail du semencier et présente de nombreux avantages, surtout dans le cadre de l'agriculture intensive :
- Amélioration du rendement : Produire davantage en moins de temps.
- Création d'espèces plus esthétiques : Par exemple, des tomates parfaitement rondes.
- Obtention de plantes plus résistantes : S'adaptant plus facilement à une production agricole intensive, aux maladies, aux parasites.
- Rapidité de croissance et goût plus prononcé.
- Homogénéité des produits : Sur le plan physique (coloris, taille) et comportemental (résistance, conservation).
Cependant, l'hybridation présente aussi des inconvénients :
- Graines inertes : À partir de la seconde génération d'hybrides, les graines des fruits ne peuvent plus germer et produire de nouvelles plantes.
- Impact sur la biodiversité : L'hybridation peut nuire à la biodiversité, d'où la nécessité de protéger les variétés sauvages en favorisant les agricultures traditionnelles et en évitant le contact des espèces sauvages avec des graines issues des premières générations d'hybrides.
Les Biotechnologies au Service de l'Amélioration des Plantes
Les biotechnologies offrent des solutions pour surmonter les limites des croisements sexués (incompatibilité, imprécision et temps) :
Culture in vitro : Permet de régénérer des plantes à partir de petits fragments de tissus (méristème) ou de cellules. Les vitro-plantes sont très répandues.
Sauvetage d'embryons immatures : En cas de barrières naturelles empêchant le développement complet de l'embryon, les embryons immatures sont prélevés à un stade précoce et mis en culture sur un milieu artificiel nutritif. Cette technique a par exemple permis d'introduire la résistance à des maladies présentes dans l'espèce sauvage Lycopersicon peruvianum chez la tomate cultivée.
Haploïdie : La régénération d'une plante à partir de cellules sexuelles (haploïdes) permet d'obtenir rapidement une plante haploïde. Le doublement du stock de chromosomes permet ensuite d'obtenir une plante diploïde, fertile et strictement homozygote. Vegenov réalise du doublement chromosomique sur les géniteurs par traitement chimique pour faciliter les croisements interspécifiques et restaurer la fertilité des hybrides interspécifiques.
Fusion de protoplastes (hybridation somatique) : Les parois cellulaires sont digérées enzymatiquement pour obtenir des protoplastes sphériques. La fusion de ces protoplastes permet d'obtenir de nouvelles plantes.
Transformation génétique : Cette technique permet d'introduire des gènes provenant de n'importe quel autre organisme. Les plantes obtenues sont dites Organismes Génétiquement Modifiés (OGM). L'obtention de la première plante transgénique en 1983 a été précédée par la découverte d'un transfert génétique naturel datant de l'antiquité. Cette méthode offre une source de gènes étendue, permettant de franchir la barrière des espèces, des genres et des règnes, et de transférer un gène précis. Les OGM permettent d'améliorer la résistance des plantes aux virus, aux insectes, aux herbicides, ou encore d'augmenter la qualité nutritive.

Les outils de base de l'amélioration des plantes sont le croisement et la sélection, afin de réunir le maximum de gènes favorables dans un même génotype. L'autofécondation et l'haplo-diploïdisation permettent de créer des lignées reproductibles. Le doublement chromosomique permet de modifier certains caractères et facilite l'échange de gènes entre espèces par la voie sexuée ; il permet aussi de créer de nouvelles espèces. Ces techniques permettent non seulement de multiplier des individus d'intérêt, mais aussi de conserver les caractéristiques. Les descendants de croisements intra et interspécifiques peuvent être caractérisés par différentes méthodes selon les besoins.
tags: #livre #les #croisements #en #horticulture