La mousse Physcomitrella patens, appartenant à la famille des Funariaceae, a émergé comme un système modèle crucial pour l'étude de l'évolution de diverses caractéristiques végétales au sein des premières lignées de plantes terrestres. En tant que première bryophyte dont le génome complet a été séquencé (Rensing et al., 2008), elle occupe une position évolutive unique à l'interface entre les algues vertes aquatiques et les plantes vasculaires. Cette position la rend particulièrement adaptée aux comparaisons phylogénétiques, offrant des aperçus précieux sur les changements génomiques associés à l'occupation de la terre. Ce guide vise à fournir des informations détaillées sur les conditions de culture et les techniques d'observation de cette mousse modèle, en s'appuyant sur des études récentes.
Contexte Évolutif et Importance de Physcomitrella patens
L'avènement de la colonisation et la diversification subséquente des premières lignées ancestrales d'Embryophyta (Kenrick et Crane, 1997; Qiu et al., 2006) ont entraîné une période d'innovations morphologiques sans précédent chez les premières plantes terrestres. Le phylum Bryophyta forme la lignée basale des plantes terrestres (Aoki et al., 2003), possédant une morphologie simple et représentant l'archétype ancestral des plantes terrestres (Renzaglia et al., 2000). Étant donné que ces bryophytes contemporaines représentent des descendants éloignés des premières plantes terrestres dont sont également dérivés les trachéophytes modernes (Hébant, 1977), ces plantes offrent une perspective unique pour examiner les premières adaptations morphologiques nécessaires à l'existence sur terre (Renzaglia et al., 2000). Parmi ces innovations morphologiques, l'évolution des tissus vasculaires est considérée comme l'un des événements les plus importants de l'histoire évolutive des plantes (Prigge et Bezanilla, 2010).
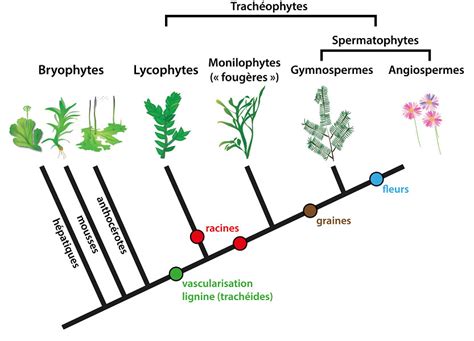
Physcomitrella patens est une plante terrestre non vasculaire, ce qui signifie qu'elle ne possède pas de vrais tissus vasculaires comme le xylème et le phloème, caractéristiques des plantes supérieures. Cependant, le gamétophore de Physcomitrella possède une anatomie de tige relativement complexe avec un brin central conducteur d'eau composé d'analogues du xylème appelés hydroids (Sakakibara et al., 2003). La présence ou non de cellules conductrices de nourriture dans la tige du gamétophore de Physcomitrella n'est pas encore totalement élucidée. Néanmoins, les mousses endohydriques comme Physcomitrella possèdent souvent des cellules conductrices de nourriture spécialisées, interprétées comme ressemblant aux cellules criblées du phloème (Behnke, 1975; Behnke and Sjolund, 2012). Ces cellules, également appelées leptoids (Hébant, 1970, 1975; Behnke, 1975), sont considérées comme des analogues des éléments criblés (Ligrone et al., 2000) et ont été postulées pour former un conduit symplasmique pour le mouvement du saccharose dans le gamétophore (Reinhart et Thomas, 1981; Raven, 2003).
Modes de Transport des Photosynthates
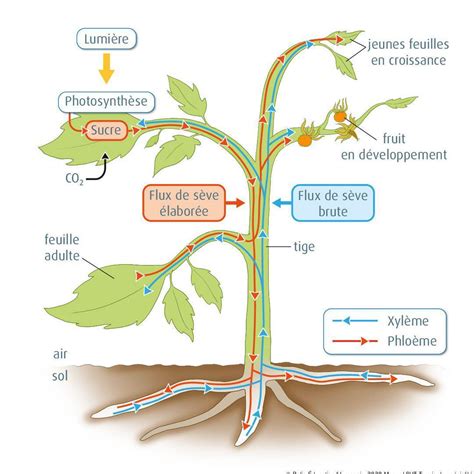
Le mode de transport des photosynthates et leur partitionnement chez Physcomitrella n'ont pas été largement rapportés. Cependant, des orthologues de transporteurs de saccharose couplés aux H+ (SUTs) (Kühn et Grof, 2010; Reinders et al., 2012a) et de protéines SWEET (Sugars Will Eventually be Exported Transporter) (Eom et al., 2015) sont présents dans le génome de Physcomitrella, suggérant l'existence de voies de transport symplasmique et apoplasmique.
Le transport symplasmique des photosynthates est prédominant dans la génération gamétophytique haploïde de la mousse. Cependant, la génération sporophytique diploïde transitoire, largement hétérotrophe, dépend nutritionnellement du gamétophyte photosynthétique (Ligrone et Gambardella, 1988; Ligrone et al., 1993). Cette dépendance fonctionnelle se reflète dans les spécialisations morphologiques des cellules de transfert situées à l'interface entre le sporophyte et le gamétophyte. Ces cellules de transfert se caractérisent par leurs invaginations étendues de la paroi cellulaire, un cytoplasme dense riche en ribosomes et des mitochondries abondantes (Pate et Gunning, 1972; Browning et Gunning, 1979a; Lal et Chauhan, 1981; Ligrone et al., 1993).
À la jonction sporophyte-gamétophyte chez Physcomitrella, une barrière de diffusion est présente, nécessitant le franchissement d'une limite apoplasmique par les métabolites (Courtice et al., 1978). Cela implique que la translocation acropète du saccharose à travers l'interface gamétophyte-sporophyte (l'haustorium) implique probablement un mécanisme de chargement apoplasmique, l'énergie métabolique pour ce mouvement directionnel du saccharose étant fournie par les nombreuses mitochondries des cellules de transfert.
Des études ont montré que les cellules de transfert des mousses sont caractérisées par une activité enzymatique intense (Hebant et Suire, 1974; Chauhan et Lal, 1982). Par exemple, Browning et Gunning (1979b,c) ont démontré, en utilisant des haustoria excisés de Funaria hygrometrica, que l'afflux de glucose et de saccharose appliqués extérieurement dans les haustoria était inhibé par des découpleurs métaboliques. À l'inverse, d'autres études ont suggéré une non-continuité du saccharose à la jonction gamétophyte-sporophyte, les hexoses étant les sucres prédominants à cette interface (Renault et al., 1992). Elles ont même avancé que la force proton-motrice établie au niveau de la membrane plasmique des cellules de transfert dans l'haustorium ne joue pas un rôle majeur dans l'absorption du saccharose, mais plutôt dans l'absorption des acides aminés (Renault et al., 1989, 1992). Leurs études physiologiques ont plutôt indiqué un mécanisme dans lequel les invertases de la paroi cellulaire hydrolysent le saccharose en hexoses, ces hexoses étant reconvertis en saccharose lors de l'absorption dans l'haustorium (Renault et al., 1992).
Cependant, des analyses enzymatiques des tissus gamétophytiques et sporophytiques de plusieurs mousses ont révélé des activités substantielles de toutes les enzymes associées à la voie de la saccharose synthase (SUS) (Galloway et Black, 1989). Compte tenu du fait que la voie de l'invertase du métabolisme du saccharose est deux fois plus coûteuse en termes de consommation d'ATP que la voie de la SUS dépendante du pyrophosphate (PPi) (Black et al., 1987), il est plausible que les mousses utilisent le mode de métabolisme de la SUS, plus économe en énergie, pour nourrir le tissu sporophytique. Cela impliquerait à son tour que l'homéostasie du PPi est cruciale pour le bon fonctionnement des cellules de transfert de mousse, de manière analogue à ce qui est observé dans le phloème des espèces à chargement apoplasmique du phloème comme Arabidopsis thaliana (Pizzio et al., 2015).
Procreation des mousses et des prêles 5min48 Y
Des investigations ultrastructurales, physiologiques, de traçage fluorescent et immunohistochimiques ont été menées pour élucider un modèle cohérent de transport des photosynthates chez Physcomitrella. Les observations ultrastructurales ont révélé que Physcomitrella est une mousse endohydrique avec des cellules conductrices d'eau et de putatives cellules conductrices de nourriture dans la tige et les feuilles gamétophytiques. Le mouvement du traceur fluorescent 5(6)-carboxyfluorescéine diacétate a montré que le mode de transport dans la génération gamétophytique est symplasmique et est médiatisé par les plasmodesmes, tandis qu'il existe une barrière de diffusion composée de cellules de transfert qui sépare le gamétophyte photoautotrophe du sporophyte hétérotrophe dépendant nutritionnellement. Il a été postulé que, de manière analogue à ce qui est trouvé chez les plantes supérieures à chargement apoplasmique du phloème, le saccharose, le principal photosynthate, est activement importé dans les cellules de transfert par des symporteurs saccharose/H+ (SUTs) qui sont, à leur tour, alimentés par des ATPases de type P, et que les cellules de transfert abritent une voie de la saccharose synthase (SUS) qui conserve l'ATP. Cette hypothèse a été étayée par la constatation qu'un protonophore (2,4-dinitrophénol) et un inhibiteur spécifique des SUT (diéthyl pyrocarbonate) réduisaient l'absorption de saccharose radiomarqué dans les sporanges. L'immunolocalisation in situ de l'ATPase de type P, de la saccharose synthase et de la pyrophosphatase protonique - tous des composants clés de la voie SUS - a montré que ces protéines étaient localisées de manière proéminente dans les cellules de transfert, fournissant des preuves supplémentaires cohérentes avec cet argument. Ces résultats suggèrent qu'il existe des modes alternatifs de transport des photosynthates chez les générations alternées de Physcomitrella, où le transport du saccharose est majoritairement passif et symplasmique chez les gamétophytes, tandis que le saccharose est activement chargé de manière apoplasmique dans les cellules de transfert via la voie SUS dans la génération sporophytique.
Conditions de Culture et Induction de la Sporophyte
Pour la culture de Physcomitrella patens, la souche sauvage Gransden est couramment utilisée.
Gamétophytes
Les gamétophytes peuvent être cultivés de deux manières principales :
- Sur milieu solide BCD en boîtes de Pétri : Cette méthode permet un contrôle précis des nutriments et facilite l'observation individuelle des cultures.
- Sur sol fertilisé par un milieu liquide BCD : Cette approche mime davantage un environnement naturel et peut être utilisée pour des cultures à plus grande échelle.
Dans les deux cas, les gamétophytes sont maintenus dans une chambre de croissance avec un régime de lumière de 16 heures et d'obscurité de 8 heures (Cove et al., 2009). Ce cycle lumineux est essentiel pour une croissance optimale et le développement photosynthétique de la mousse.
Sporophytes
L'induction de la croissance des sporophytes est un processus spécifique qui nécessite des conditions environnementales particulières. Les gamétophytes matures âgés de 4 à 5 semaines sont transférés dans une chambre maintenue à 15°C sous un régime de 8 heures de lumière et 16 heures d'obscurité. Ces conditions sont optimisées pour déclencher la reproduction sexuée et le développement du sporophyte, qui est la génération diploïde de la mousse et est nutritionnellement dépendante du gamétophyte.
Techniques d'Observation et d'Analyse
Diverses techniques sont employées pour étudier la morphologie, l'ultrastructure et les mécanismes physiologiques de Physcomitrella patens.
Microscopie Optique
La microscopie optique est une technique fondamentale pour l'observation des gamétophytes matures.
- Stéréomicroscope Olympus SZX7 : Cet équipement, couplé à un système de caméra Infinity 2 et au logiciel d'imagerie Infinity Capture (Lumenera Corporation), permet d'imager des gamétophytes individuels avec une grande clarté.
- Microscope inversé Nikon Eclipse TE300 : Pour des observations plus détaillées, notamment de sections semi-fines (250-500 nm) de tiges noyées dans de la résine, la microscopie optique est utilisée. Ces sections sont colorées au bleu de toluidine, ce qui permet de visualiser les structures cellulaires. L'équipement est généralement muni d'une caméra Olympus DP26 et du logiciel d'imagerie numérique cellSens (Olympus Corporation).
Microscopie Électronique à Transmission (MET)
La MET fournit des images à très haute résolution des structures internes des cellules.
- Préparation des échantillons : Des gamétophores matures individuels de Physcomitrella sont fixés dans une solution de glutaraldéhyde à 4,0 % et de formaldéhyde à 1,0 % dans un tampon phosphate de sodium 0,05 M, pH 7,2 pendant 2 heures à température ambiante. Les feuilles sont délicatement séparées des tiges sous stéréomicroscope. Les tiges feuillées et nues sont ensuite découpées en morceaux d'environ 2 mm.
- Post-fixation et coloration : Les tissus sont ensuite post-fixés dans de l'osmium tétroxyde (OsO4) à 1 % pendant 2 heures à température ambiante, puis colorés en bloc avec de l'acétate d'uranyle à 0,2 % pendant une nuit à 4°C.
- Déshydratation et inclusion : Les tissus sont ensuite déshydratés en série dans de l'acétone et progressivement infiltrés avec de la résine de Spurr sur 5 à 7 jours. Ils sont ensuite noyés à plat dans des moules ou sur des lames de verre revêtues de Téflon dans un four à 60°C. Les tissus sont noyés dans diverses orientations pour faciliter la coupe longitudinale et transversale. Tous les produits chimiques de fixation sont généralement achetés auprès d'Electron Microscopy Sciences (Washington, PA, États-Unis).
Microscopie Électronique à Balayage (MEB)
La MEB permet d'obtenir des images de surface détaillées des gamétophores et sporophytes.
- Fixation et déshydratation : Des gamétophores et sporophytes individuels sont fixés avec du glutaraldéhyde à 4 % dans un tampon phosphate de sodium 0,05 M, pH 7,2 pendant 4 à 6 heures à température ambiante, post-fixés dans de l'OsO4 à 1 % pendant 2 heures à température ambiante, déshydratés en série avec de l'acétone, et séchés au point critique (Balzers Union, Principauté de Liechtenstein).
- Dissection et traitement : Si les tissus doivent être disséqués, les échantillons séchés au point critique sont coupés de manière appropriée à l'aide d'une lame de rasoir fraîche à double tranchant sous un stéréomicroscope. Les produits chimiques de fixation sont généralement achetés auprès d'Electron Microscopy Sciences.
- Traitement d'image : Les micrographies électroniques à balayage et à transmission (MEB et MET, respectivement) sont ensuite traitées à l'aide du logiciel Adobe Photoshop CC (Adobe Systems Incorporated). Des ajustements linéaires (luminosité, contraste et niveaux) sont effectués sur les images entières pour améliorer la clarté.
Étude des Traceurs Fluorescents
Cette méthode permet de suivre le mouvement des substances à travers les tissus.
- Application du traceur : Des gamétophores individuels ou des gamétophores porteurs de sporophytes sont verticalement flottés sur environ 5 μL d'une solution de 5(6)-carboxyfluorescéine diacétate (CFDA) ou de HPTS à 250 μg.mL-1 dans des barrettes de tubes PCR.
- Incubation : Les échantillons sont ensuite incubés pendant 1, 3 ou 24 heures dans une chambre humide sombre. Des contrôles négatifs sont effectués en faisant flotter les plantes uniquement dans de l'eau.
- Préparation des sporophytes : Les gamétophores porteurs de sporophytes sont traités de manière identique, à l'exception de l'excision de la plupart des feuilles avant l'imagerie.
- Étude sur feuilles excisées : Pour l'étude du traceur CFDA dans des feuilles excisées individuellement, des feuilles uniques sont soigneusement retirées de la tige à l'aide de fines pinces sous un stéréomicroscope et traitées de manière identique, sauf qu'uniquement 1 à 2 μL de solution de CFDA sont utilisés.
Coloration au Bleu d'Aniline et Microscopie Confocale
Cette technique est utilisée pour visualiser les plasmodesmes, qui sont des canaux intercellulaires.
- Procédure de coloration : La coloration au bleu d'aniline de la callose associée aux plasmodesmes des feuilles de Physcomitrella est réalisée selon le protocole décrit par Zavaliev et Epel (2015), en utilisant 0,01 % (p/v) de bleu d'aniline dans du K3PO4 0,01 M, pH 12.
- Montage et imagerie : Les feuilles sont ensuite montées entières dans la solution de coloration elle-même et imagées avec un microscope confocal spectral Leica TCS SP5 AOBS (Leica Microsystems) équipé d'un objectif sec Plan-Apo 20×/0,7 NA, d'optiques DIC et du logiciel confocal Leica. La coloration au bleu d'aniline associée aux plasmodesmes est imagée (512 pixels × 512 pixels) à une longueur d'onde d'excitation de 405 nm, et le signal d'émission est détecté entre 475 et 525 nm pour le bleu d'aniline, et entre 643 et 730 nm pour l'autofluorescence de la chlorophylle. Des contrôles négatifs sont réalisés en excluant le bleu d'aniline.
Immunohistochimie
L'immunohistochimie permet de localiser spécifiquement des protéines clés au sein des tissus.
- Fixation des échantillons : Des gamétophores entiers et des gamétophores porteurs de sporanges sont immédiatement immergés dans un volume au moins 20 fois supérieur de FAA (formaldéhyde à 10 % v/v 37 % : acétone à 5 % v/v…). Cette méthode est cruciale pour étudier la localisation de protéines telles que les SUTs, les ATPases de type P, les H+-PPases et les SUS, qui sont impliquées dans le transport des photosynthates.
Conséquences des Études sur le Repiquage
La compréhension approfondie des mécanismes de transport des photosynthates et des conditions de croissance optimales pour Physcomitrella patens a des implications directes pour le repiquage. Un repiquage réussi dépend de la capacité de la plante à s'établir et à maintenir un transport efficace des nutriments.
- Choix du stade de développement : Le repiquage des gamétophytes matures, plutôt que des stades immatures, peut améliorer le taux de survie, car ils ont déjà établi des structures de support et un système de transport des photosynthates plus robuste.
- Conditions environnementales post-repiquage : Le maintien des conditions lumineuses et de température spécifiques au gamétophyte (16h lumière/8h obscurité) est essentiel pour éviter le stress. Lors de l'induction du sporophyte, le passage à 15°C et un régime de 8h lumière/16h obscurité doit être respecté pour une transition réussie.
- Intégrité des tissus : Lors du repiquage, il est crucial de minimiser les dommages aux tissus, en particulier aux tiges et aux feuilles, où les systèmes de transport d'eau et de photosynthates sont localisés. Les dommages pourraient perturber le mouvement symplasmique et apoplasmique des sucres, ralentissant la récupération de la plante.
- Milieu de culture : L'utilisation d'un milieu BCD, qu'il soit solide ou liquide, comme décrit dans les méthodes de culture, est fondamentale pour fournir les nutriments nécessaires. Un sol bien fertilisé ou un milieu solide correctement préparé sont des facteurs clés.
- Minimiser les contraintes : La dépendance nutritionnelle du sporophyte envers le gamétophyte souligne l'importance de maintenir un gamétophyte sain lors du repiquage des sporophytes. Toute perturbation de la jonction gamétophyte-sporophyte pourrait avoir des conséquences néfastes sur la croissance du sporophyte.
En respectant ces considérations basées sur la physiologie et l'ultrastructure de Physcomitrella patens, les protocoles de repiquage peuvent être optimisés pour assurer la viabilité et la croissance continue de cette mousse modèle précieuse pour la recherche.
tags: #moss #physcomitrella #repiquage