La compréhension du vivant repose sur deux piliers fondamentaux : la capacité humaine à manipuler les outils biologiques pour optimiser les performances des populations animales, et la connaissance des mécanismes naturels qui régissent l’évolution des espèces sur le temps long. Alors que la semence sexée permet une gestion dirigée et immédiate des cheptels, les forces évolutives comme la dérive génétique dictent, de manière stochastique, le destin génétique des populations.
La technologie de la semence sexée en élevage
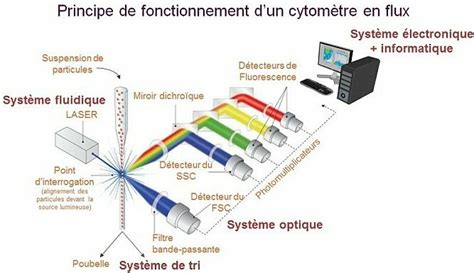
La semence sexée vient d’une méthode inventée dans les années 1980 et qui a été brevetée et mise en place par la firme américaine Sexing Technology en 2003. Elle est arrivée dans les coopératives d’insémination dans les années 2009. Le tri se fait par cytométrie de flux. Le principe est simple, la différence entre un spermatozoïde mâle et femelle est le chromosome X et Y distribué aléatoirement après la méiose. Le tri des spermatozoïdes se fait simplement par rapport au poids.
L’utilisation de la semence sexée permet à l’éleveur d’avoir le choix et il peut s’avérer très important ! Que ce soit pour l’éleveur qui cherche à performer en génétique, celui qui souhaite avoir un bon renouvellement ou encore pour valoriser mieux les produits pour la vente. Aujourd’hui l’utilisation de la semence sexée représente 429 581 IA en France en 2020. Pour garder un nombre suffisant de femelles en élevage notamment laitier pour la production ou pour augmenter son cheptel, le nombre de veaux femelles par an est important. C’est là que la semence sexée est un outil pratique pour gérer la variabilité du sexe et truquer les dés en se sécurisant un nombre de femelles sûr.
La semence sexée permet d’être sûr d’avoir une descendance femelle pour garder la génétique d’une vache que l’on souhaite conserver et améliorer. Ainsi en mettant un bon taureau sexé avec une bonne femelle, la descendance femelle sera toujours présente au sein du troupeau. La valorisation des veaux en fonction des filières est très disparate. Par exemple des veaux mâles engraissés ont des prix de vente souvent plus élevés surtout en élevage allaitant. La semence sexée mâle permet d’obtenir des produits mieux valorisés à la vente. Quant à la semence sexée femelle, elle permet de vendre des femelles pleines mieux valorisées.
Défis techniques et optimisation de la fertilité
La semence sexée reste des doses bien plus chères que la semence classique, et elle est aussi moins fertile dû au traitement de la semence qu’elle subit et du fait que la concentration d’une paillette sexée est moindre qu’une paillette conventionnelle : 2,1 millions de spermatozoïdes contre 18 millions. Pour l’instant les outils ne permettent pas de rendre plus fiable la semence. Le pourcentage de chance d’obtenir le sexe voulu, mâle ou femelle, est de 90%.
Pour pallier à tout cela, il faut être vigilant sur sa repro et échanger avec un expert (vétérinaire, inséminateur, etc…) pour inséminer en optimisant la reproduction. Le planning d’accouplement est aussi un bon outil à coupler avec la semence sexée pour optimiser et bien cibler les femelles choisies. Enfin pour réduire la perte de fertilité, il faut favoriser l’utilisation sur génisse qui a un meilleur taux de fertilité. Sinon il existe des outils comme XtremiA permettant d’amener au bout de la corne le faible nombre de spermatozoïdes et donc d’éviter cette perte de charge le long de la remontée de l’utérus.
IVI Fertilité - Insémination artificielle (UE, 2014)
Les fondements biologiques de l’évolution : mutations et génomes
Toutes les espèces présentes à l’heure actuelle sur notre planète sont caractérisées par une information génétique, contenue dans leur génome, qui leur est propre. Cela dit, le génome de ces espèces n’est pas figé. Divers mécanismes comme les duplications, les fusions ou les cassures de chromosomes, les mutations géniques (insertions, délétions, substitutions…) contribuent à faire évoluer ces génomes, conduisant, à terme, à partir d’une espèce ancestrale, à l’apparition de groupes d’individus qui, du fait des nombreuses modifications au sein de leur génome, ne sont plus interféconds et constituent donc de nouvelles espèces.
S’il y a non-disjonction des chromosomes de la lignée germinale en méiose, cela va permettre la création de gamètes diploïdes à 2n chromosomes (au lieu de n normalement). S’il y a fusion entre deux gamètes de ce type, on va obtenir un zygote à 4n chromosomes qui aura donc subi un doublement complet de son génome. Lors de l’hybridation interspécifique entre 2 espèces donnant 2 lots chromosomiques distincts, le nombre de chromosomes de l’hybride F1 est alors la somme des chromosomes de chaque espèce. Le "triangle de U" décrit les relations génétiques entre 3 espèces de Brassica diploïdes (B. oleracea, B. rapa et B. nigra) et 3 espèces allotétraploïdes (B. napus, B. juncea et B. carinata).
Les gènes présents au sein des génomes peuvent évoluer, se modifier du fait des forces évolutives par des mécanismes aléatoires (mutations, recombinaisons…) et d’autres non aléatoires (sélection naturelle sexuelle, sélection artificielle…). Les différents allèles d’un gène n’apparaissent pas en même temps durant l’évolution d’une espèce. De plus, ils peuvent conférer un avantage ou un désavantage aux individus qui les portent. De manière générale, l’ensemble des mécanismes décrits comme étant des forces évolutives (sélection naturelle, mutation, recombinaison, dérive, migration…) contribue à faire varier les fréquences alléliques des gènes.
Le mécanisme de la dérive génétique
La dérive génétique concerne l’évolution au sein d’une population, ou d’une espèce, de la fréquence des allèles ou des génotypes (combinaison des deux allèles d’un gène héritée des chromosomes paternel et maternel au moment de la fécondation) d’un gène, causée par des phénomènes aléatoires et impossibles à prévoir, donc indépendamment des mutations, de la sélection naturelle et des migrations. Les effets de la dérive génétique sont d’autant plus importants que la population est petite, car les écarts observés d’une génération à l’autre par rapport aux fréquences alléliques y sont d’autant plus perceptibles.
La dérive génétique concerne surtout les allèles neutres, qui ne confèrent ni avantage ni désavantage sélectif. Alors que dans des populations de taille infinie, en l’absence de sélection et de mutation, les fréquences alléliques sont stables au cours des générations (principe de Hardy-Weinberg), elles vont varier aléatoirement dans des populations diploïdes de taille finie. Lors de la fécondation, la rencontre de deux gamètes haploïdes paternel et maternel va donner naissance à un nouvel individu diploïde. Si pour un gène donné les deux copies provenant des deux parents sont identiques, on dit que l’individu est homozygote. C’est lors de la séparation des chromosomes homologues qu’il va y avoir une répartition, un tirage, aléatoire des chromosomes allant dans chaque gamète.
A l’échelle d’une population, il est donc statistiquement inévitable que certains allèles ne soient pas transmis d’une génération à l’autre. La dérive génétique est donc un mécanisme aléatoire (stochastique) qui va modifier la fréquence des allèles d’un gène dans la population en l’augmentant ou en la diminuant. En fonction de la taille de la population et de la fréquence des allèles à l’origine, le temps de fixation va varier. L’effet de taille de la population est particulièrement visible lorsqu’un faible groupe d’individus d’une espèce va coloniser un autre milieu. Plus la taille de la population est faible, plus la dérive est rapide.
L'effet fondateur et la consanguinité
Lorsqu’une sous-partie d’une population se sépare de la population initiale de taille beaucoup plus vaste, lors d’une migration pour coloniser un nouveau milieu par exemple, la population pionnière, ou fondatrice, n’est pas le reflet exact de la population de départ. Cette sous-population ne va « prendre » qu’un échantillon du pool d’allèles disponible dans la population mère « dans ses bagages » et ce, de manière aléatoire. Elle peut donc avoir des fréquences alléliques fort différentes de la population initiale. C’est ce que l’on appelle « l’effet fondateur ».
La consanguinité va intervenir sur les fréquences alléliques des gènes en amplifiant le taux d’homozygotie. En construisant les arbres généalogiques des familles dans lesquelles sévissent à l’heure actuelle ces maladies, on se rend compte que pour chacune il existe un ancêtre fondateur unique qui a introduit la mutation délétère. C’est uniquement par hasard que des porteurs de ces allèles délétères se sont trouvés dans la population fondatrice. Par la suite le mode de reproduction en milieu confiné, accroissant la consanguinité, a amplifié la fréquence de ces allèles délétères dans les familles. Les études de fixation d’allèles par dérive génétique ont montré que ce mécanisme peut aboutir à la fixation d’un allèle du gène au détriment des autres, dans une population.
Sélection naturelle et sélection sexuelle : les forces orientées
La sélection naturelle est le processus par lequel les organismes mieux adaptés à leur environnement tendent à survivre et à se reproduire plus que ceux moins adaptés. Elle agit sur tous les types d’organismes vivants et s’opère au niveau des gènes. Elle favorise les adaptations avantageuses, qui peuvent être des caractères physiques ou comportementaux, en augmentant la fréquence des allèles bénéfiques dans la population.
La sélection sexuelle est un concept introduit par Darwin pour expliquer certaines caractéristiques qui ne semblent pas avoir de rôle spécifique dans la survie des organismes, comme les grandes plumes du paon ou le bois des cerfs. Ces caractéristiques peuvent néanmoins offrir un avantage reproductif en rendant un individu plus attrayant aux yeux de ses partenaires potentiels. Deux principaux mécanismes de sélection sexuelle existent : la compétition intrasexuelle, où les membres d’un sexe rivalisent pour l’accès aux partenaires, et la sélection intersexuelle, où un sexe choisit des partenaires basés sur certains traits attractifs.
La complexité du génome et les mécanismes de variabilité
Le code génétique est redondant : certains acides aminés peuvent être codés par plusieurs codons différents. Lorsque la mutation est non synonyme, cela ne signifie pas pour autant que le fonctionnement de la protéine est modifié. Souvent, seules certaines parties de la molécule ne supportent aucun changement, tandis que les autres parties peuvent supporter le remplacement d’un acide aminé par un autre, en général à condition que les propriétés physiques et chimiques de celui-ci ne soient pas trop différentes de l’état originel.
Une part appréciable des génomes est composée d’éléments mobiles (ou éléments transposables). Certains se déplacent par un mécanisme d’excision/réinsertion (éléments à ADN), tandis que d’autres sont transcrits en ARN, puis réverse-transcrits en ADN, la copie étant ensuite réinsérée ailleurs dans le génome (rétroéléments). Ces séquences sont des sites qui permettent la fixation de protéines spécifiques, qui peuvent bloquer ou au contraire permettre la transcription du gène. De nombreux chercheurs pensent que ce type de modifications représente l’une des clés pour comprendre la diversité de forme des organismes.
Reproduction sexuée et brassage génétique
Les cellules reproductrices (= gamètes), possèdent 23 chromosomes dans leur noyau. Lors de la formation des gamètes, on observe une réduction chromosomique lors de la méiose. Un des deux chromosomes de chaque paire de chromosomes contenu dans les cellules des testicules ou des ovaires se retrouve dans les cellules reproductrices. En réunissant les 23 chromosomes du spermatozoïde et les 23 chromosomes de l’ovule, la fécondation aboutit à une cellule-œuf avec 46 chromosomes.
Lors de la formation des gamètes, la répartition des chromosomes s’effectue au HASARD pour chaque paire. La méiose et la fécondation permettent la formation d’une cellule-œuf qui sera à l’origine d’un nouvel individu original et génétiquement unique. La reproduction sexuée aboutit à des individus tous différents, à l’exception des vrais jumeaux. Les mutations génétiques sont des changements qui interviennent au niveau de l’ADN. Ces changements peuvent induire de nouveaux allèles et donc de nouveaux caractères héréditaires.
Synthèse des forces évolutives
Les importances relatives de la sélection, de la dérive et de la mutation dans le processus évolutif sont encore discutées. Il faut également garder à l’esprit qu’un individu est une combinaison de très nombreux allèles, et que les effets de divers allèles sont parfois difficilement séparables. Il peut sembler d’après ces descriptions que l’évolution est entièrement aléatoire. Cela n’est pas totalement le cas.
La production de variabilité ainsi qu’une partie du tri de cette variabilité sont aléatoires, mais opèrent en général par modification de séquences/structures existantes : « le bricolage » décrit par François Jacob dans Le Jeu des Possibles. A un moment donné, dans un individu donné, tout n’est donc pas possible. Ce concept de réutilisation et modification de structures pour créer de la nouveauté fonctionne à l’échelle des organes comme à celle des gènes. La sélection ramène également une certaine directionnalité ; de nouveau, tout n’est pas possible, et les porteurs de certains allèles seront plus ou moins avantagés ou désavantagés par rapport au reste de la population selon la valeur sélective des allèles les uns par rapport aux autres. Mais cette directionalité même si parfois très forte, n’est pas absolue.
La compréhension de ces mécanismes, qu'il s'agisse de la gestion pratique de la semence sexée pour orienter la descendance d'un troupeau ou de l'étude théorique de la dérive génétique, souligne la complexité permanente du vivant. Entre l'intervention humaine ciblée et les forces stochastiques naturelles, l'évolution demeure une dynamique constante de modification des fréquences alléliques, façonnant la diversité biologique à travers les générations.