Le cœur humain, dans sa complexité anatomique actuelle, est le résultat d'un processus dynamique fascinant qui prend racine dès les premières semaines de la vie embryonnaire. Pour comprendre la structure et la fonction des grandes voies de sortie du cœur, il est indispensable d'étudier le bulbe artériel, une structure clé du développement cardiaque qui assure la transition entre le ventricule primitif et les arcs aortiques.
Origine embryonnaire et mise en place du tube cardiaque
Les grandes lignes du développement du cœur humain sont utiles à connaître, ne serait-ce que pour expliquer la genèse de certaines malformations cardiaques reconnues dès la naissance, ou pendant la croissance. Les premières ébauches du système cardio-vasculaire apparaissent dès le 17ème jour, au niveau du mésoderme extra-embryonnaire du sac vitellin. À ce stade, le système s'organise de façon paire : un tube cardiaque et une aorte dorsale de chaque côté.
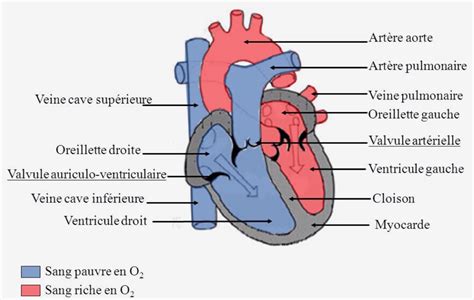
La première ébauche aligne quatre cavités, dans l'ordre : le sinus veineux, l'oreillette, le ventricule, le bulbe artériel. À la suite de coudures et d'inégalités de développement, le sinus veineux vient se placer sur le versant droit de la cavité auriculaire, et le bulbe artériel sur le versant céphalique de la cavité ventriculaire. Vers la 23ème journée, la croissance inégale du tube cardiaque provoque un repliement spécifique. Ce processus dynamique positionne l’oreillette primitive et les raccords veineux derrière le ventricule primitif et le bulbe artériel. Ce repliement est essentiel pour organiser les futures cavités cardiaques et préparer la différenciation des ventricules.
La spécialisation du bulbe artériel (Bulbus Cordis)
Une structure comparable au bulbe artériel (bulbus arteriosus) est traversée pendant les phases de développement du cœur humain et est également appelée bulbus cordis. Celui-ci se trouve ventral au ventricule primitif après que le cœur en développement a pris sa forme en S. Ensemble, le bulbus cordis et le ventricule primitif forment les ventricules du cœur ainsi formés.
Les parois adjacentes du bulbus cordis et du ventricule s'approchent, fusionnent et finissent par disparaître de sorte que le bulbus cordis communique maintenant librement avec le ventricule droit, tandis que la jonction du bulbe avec le tronc artériel est amenée ventralement dans le canal auriculaire. Le bulbe artériel subit de même un cloisonnement par le septum aorticum ou septum bulbi, qui a la particularité de décrire un trajet spiral ; la partie supérieure, orientée d'avant en arrière, se raccorde insensiblement avec la partie inférieure, dirigée transversalement, de gauche à droite.
Cloisonnement aortico-pulmonaire et devenir des vaisseaux
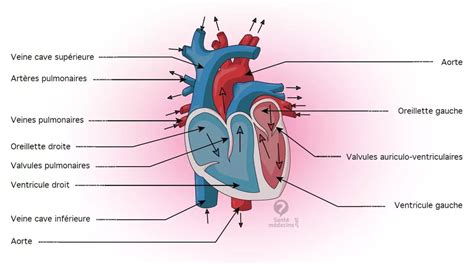
Le cloisonnement du bulbe artériel est un mécanisme distinct de la séparation linéaire observée dans les oreillettes et les ventricules. Ce processus suit une structure en spirale appelée cloisonnement aortico-pulmonaire. Cette division rencontre tout d'abord les valvules latérales du bulbe artériel primitif, à leur partie moyenne. Ce faisant, elle isole deux vaisseaux, de calibre comparable, l’artère pulmonaire en avant, munie de la valvule ventrale du bulbe, et l’aorte en arrière, munie de la valvule dorsale.
Poursuivant sa progression dans la cavité ventriculaire, le septum aorticum rejoint le septum inferius, ferme le trou de Panizza, et achève ainsi le cloisonnement du ventricule primitif en ventricule droit et ventricule gauche. L'ouverture persistante entre les deux ventricules, présente au sommet du septum musculaire, se ferme grâce à l’incorporation d’une partie du bulbe artériel. Elle joue un rôle crucial en scellant la communication entre les ventricules.
Perspectives comparatives : le bulbe artériel chez les vertébrés inférieurs
Dans le système circulatoire des poissons, le bulbe artériel est une chambre en forme de poire qui fonctionne comme un condensateur, maintenant un flux sanguin continu dans les arcs branchiaux. Le bulbe artériel distingue la chambre la plus antérieure du cœur des Téléostéens et des cyclostomes, un bulbe sans valvules et non contractile à la différence de son équivalent chez les autres poissons, le bulbe cardiaque.
Le sang est envoyé par le cœur aux artères qui irriguent les différentes branchies : ce sont les artères branchiales afférentes. Le sang repart des branchies par des artères branchiales efférentes qui vont distribuer le sang oxygéné aux organes. On peut remarquer que le cœur n’est pas double comme chez les mammifères (pas de cœur droit veineux pas de cœur gauche artériel), il est uniquement veineux. Le sang arrive par le sinus veineux, passe par une cavité appelée atrium (équivalent de l’oreillette) puis pénètre dans le ventricule. Lorsque le ventricule se contracte, il envoie le sang dans le bulbe artériel puis dans l’aorte ventrale.
Le fonctionnement du cœur expliqué par Jamy
Implications cliniques et malformations cardiaques
Ce développement harmonieux et achevé du septum inferius et du septum bulbi n’est pas toujours réalisé. Il en résulte différentes malformations. La persistance du trou de Panizza entraîne une communication interventriculaire, avec hyperpression au niveau du ventricule droit. Elle peut s'associer à une dextroposition de l'aorte, qui se place à cheval sur les deux cavités ventriculaires.
La non-occlusion du trou de Botal crée également une malformation : la communication interauriculaire. Ce peut être simplement une petite fistule, mais aussi ce peut être une vaste brèche, par agénésie du septum primum. Ces conditions démontrent l'importance vitale du timing embryologique dans la fusion des cloisons, le septum primum et le septum secundum, ainsi que la progression du septum aorticum.
Organisation vasculaire et terminaisons carotidiennes
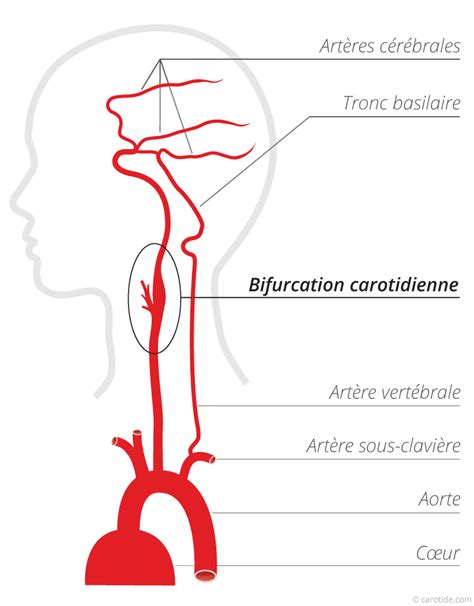
Le bulbe artériel joue un rôle précurseur dans la structuration des grands troncs artériels. La vascularisation artérielle du cerveau est assurée par quatre artères qui irriguent les différents territoires vasculaires cérébraux : les deux artères carotides en avant et les deux artères vertébrales en arrière. La carotide commune gauche naît de l'aorte et se divise dans le cou en carotide interne vers le cerveau et carotide externe vers le visage et le cou. Elle naît du TABC (tronc artériel brachio-céphalique) à droite et de l'aorte à gauche.
Son diamètre moyen est de 9 mm. Elle comprend plusieurs segments : le premier segment est fusiforme et s'appelle le bulbe carotidien. Il mesure 1 à 2 cms de long et son diamètre moyen est de 9 mm. C'est là que se forment la majorité des plaques d'athérome. Le second segment est la carotide interne sus-bulbaire dont le diamètre moyen est de 5 mm, puis viennent le segment sous pétreux avant l'entrée de la carotide dans la base du crâne, le segment intra pétreux situé dans l'os du crâne, le segment intra caverneux ou siphon carotidien, et le segment terminal qui se divise en 2 artères cérébrales : l'artère cérébrale antérieure et l'artère cérébrale moyenne (artère sylvienne).
Structure et histologie cardiaque lors du développement
Le cœur, en tant qu'organe principal du système circulatoire sanguin, voit ses mécanismes physiologiques s'illustrer efficacement sur des schémas simples (le cœur vu comme un rectangle divisé en quatre, étages auriculaire et ventriculaire, parties droite et gauche) qui ne rendent cependant pas compte de la complexité anatomique. Celle-ci s'aborde plus facilement en évoquant le film de la formation embryologique du système cardio-vasculaire.
Lors de la plicature de l’embryon, le sac vitellin est « pincé » pour donner l’ébauche tubulaire digestive, entourée d’une cavité creusée dans le mésoderme, qui se met en communication avec le cœlome péricardique par les canaux péricardo-péritonéaux. Parallèlement au cloisonnement du canal auriculo-ventriculaire, un mécanisme différent s’opère dans le bulbe artériel. Les ventricules poursuivent leur développement interne, où les faisceaux musculaires spongieux se réorganisent progressivement. Certains s’amincissent, tandis que d’autres se transforment en structures essentielles : les muscles papillaires et les cordages tendineux. Ces cordages, reliés aux muscles papillaires, jouent un rôle clé en maintenant les feuillets des valves auriculo-ventriculaires en position, empêchant ainsi le reflux du sang lors de la systole ventriculaire.