Le bulletin "Voyages au pays des semences" de Catherine Guy, diffusé notamment via LinkedIn, offre une plongée approfondie dans le monde complexe de la semence, en mettant en lumière des perspectives souvent négligées et en remettant en question les approches conventionnelles. À travers ce dossier technique, l'auteure explore l'histoire de la sélection du blé, les enjeux des organismes génétiquement modifiés (OGM) et les promesses de la sélection paysanne participative. L'œuvre, rédigée avant 2000, fut initialement l'un des rares textes consacrés aux céréales et à la manipulation génétique, fruit d'une bibliographie abondante. Cette époque, caractérisée par une forte opposition aux OGM, a vu les risques évidents de pollution génétique et l'accaparement du marché de la semence par formatage devenir des sujets de préoccupation majeurs. L'arrachage de plantes OGM par les faucheurs volontaires soulevait des questions fondamentales sur la légitimité des différentes parties prenantes.
Initialement, la réflexion semencière était principalement axée sur le principe de précaution face à l'intervention dans les gènes, sans pour autant proposer une alternative technique de sélection. Cependant, une rencontre fortuite avec Jean-François Berthellot, un agriculteur biologique engagé dans la réappropriation de la sélection paysanne, a transformé cette perspective. Il a remis en question les conclusions initiales de Catherine Guy, l'incitant à approfondir sa compréhension et à rencontrer des acteurs de terrain. Cette démarche a révélé l'importance de l'observation, de l'expérimentation et de la revisite des méthodes de sélection, des pratiques que Jean-François Berthellot menait avec une détermination de pionnier, sans craindre les échecs.
La Sélection Paysanne et Participative : Une Dynamique Enrichissante
La pratique de la sélection paysanne et participative, telle qu'exemplifiée par Jean-François Berthellot, ne visait pas un bénéfice personnel, mais plutôt la promotion d'une approche collective. L'aide apportée lors de la récolte de sa remarquable collection d'anciens blés devenait un moment privilégié de rassemblement sur le terrain et d'action collective. Cette rencontre s'est avérée être une expérience merveilleuse, invitant à questionner les pratiques conventionnelles qui, selon Berthellot, conduisaient à un appauvrissement continu de la diversité. Ces pratiques s'inféodaient à une prétendue "phytothérapie", diminuaient la matière nutritive en "nanifiant" les tiges et réduisaient les potentialités de formation de bonnes mycorhizes.
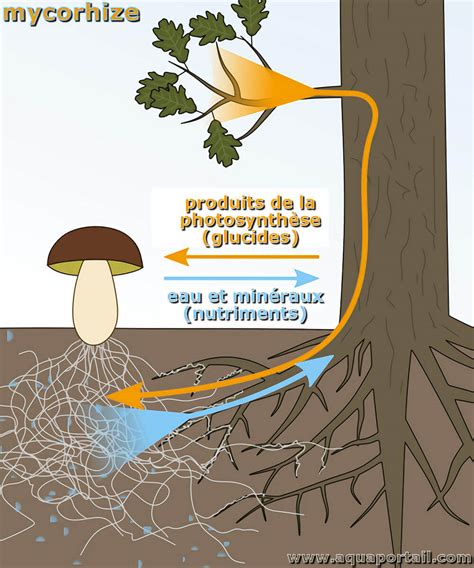
Le Rôle Crucial des Mycorhizes
Les mycorhizes représentent une symbiose remarquable entre les racines d'une plante, comme le blé, et les micro-champignons qui les entourent de leur mycélium. Cette collaboration offre de multiples avantages à la plante, la rendant plus résistante aux maladies et aux aléas climatiques. Elles facilitent également l'absorption de l'eau et de divers nutriments que ces champignons captent plus efficacement. La compréhension de cette symbiose naturelle souligne l'importance d'une approche holistique de l'agriculture, qui respecte et favorise les processus biologiques.
L'Expérience Gustative et la Quête d'un Renouveau
L'observation d'anciennes variétés, leur dégustation au moment de la récolte, leur mouture sans additifs, et leur panification par auto-fermentation, jusqu'à la décision finale du pain mis sur la table, constituaient une expérience merveilleuse. L'impatience d'apprécier le résultat le lendemain était palpable. Les premiers pains de touselle, de petit épeautre, ou de khorasan, ont procuré un bonheur inattendu, révélant une qualité gustative insoupçonnée. Cette découverte a ouvert un horizon nouveau pour tous les participants, marquant le début d'une longue amitié et, plus important encore, d'une aventure semencière et boulangère. Cette quête d'un renouveau de la sélection paysanne sera décrite plus en détail ultérieurement.
Les semences paysannes et la sélection participative
Les Graines de Semences : Une Histoire d'Amélioration et de Controverses
L'amélioration de la sélection des semences de froment et d'autres céréales panifiables a probablement été initiée par les premiers agriculteurs, qui ont abandonné la vie nomade. Ils prélevaient des graines pour leurs futures récoltes en sélectionnant simplement les plus gros grains issus des plus beaux épis. Ce type de réemploi des graines en semences et cette sélection dite « massale » sont souvent critiqués par les experts autoproclamés du secteur semencier.
L'Autogamie du Froment : Un Facteur de Stabilité
Le froment est récalcitrant à la transformation et aux manipulations génétiques en raison de son caractère autogame et de sa tendance à l'homozygotie. En termes plus simples, le froment s'autoféconde, ce qui signifie que les étamines d'une fleur fécondent presque toujours les ovaires de la même fleur. Ce phénomène se produit à l'intérieur de la balle, l'enveloppe de la graine. Le caractère autogame du froment se traduit par la reproduction du même caractère dans sa descendance, ce qui tend à le rendre homozygote.
De plus, le froment est une monocotylédone, une caractéristique qui a longtemps constitué un obstacle à la manipulation génétique. Les premiers transferts de gènes, réalisés à l'aide d'agrobactéries à partir de 1997, étaient inopérants sur les monocotylédones.
L'Émergence de CRISPR-Cas9 : Un « Ciseau Génétique »
De nos jours, des techniques telles que CRISPR-Cas9 sont employées. Il s'agit d'un "ciseau génétique" qui coupe l'ADN à l'aide d'enzymes de restriction. L'action d'une enzyme ligase ressoude ensuite les chaînes d'ADN séparées, après avoir intégré le gène d'intérêt ou retiré un gène indésirable. La Française Emmanuelle Charpentier et l'Américaine Jennifer A. Doudna ont reçu le prix Nobel de chimie en 2020 pour avoir décrit cet outil en 2012. Jennifer A. Doudna a d'ailleurs publié un livre au titre évocateur : "Un coup de ciseaux dans la Création. Le redoutable pouvoir de contrôler l'évolution". Ce type d'outil au sein du vivant nécessite un cadre légal clairement défini pour éviter tout dérapage de la recherche et empêcher que la descendance ne soit arbitrairement déclarée parfaite.
Actuellement, ces techniques sont parfois désignées comme de "Nouvelles Biotechnologies" ou OGM cachés, car elles ne sont pas toujours déclarées comme telles. Les débats s'annoncent serrés à l'avenir, alors que certains souhaitent changer la dénomination de "modification génétique" en "édition génétique".
L'Allogamie du Blé : Une Rareté Naturelle
Plusieurs chiffres circulent sur les possibilités d'allogamie du blé, c'est-à-dire de fécondation croisée par des pollens provenant d'autres variétés de Triticum, qui peut atteindre jusqu'à 10 % certaines années. Cela signifie qu'il y a 90 % de chances que ce phénomène ne se produise pas. Une fleur de blé peut rester ouverte de huit minutes à une heure. Les étamines des anthères de blé produisent en moyenne 450 000 grains de pollen. En comparaison, et pour mieux comprendre l'autogamie ou autofécondation du blé par rapport aux autres céréales, le seigle produit en moyenne 4 000 000 grains de pollen et le maïs 25 000 000 grains, soit respectivement 9 à 55 fois plus de pollen. Les grains de pollen de blé ne sont viables que 15 à 30 minutes. Du côté de la fleur hôte, le stigmate reste réceptif de 6 à 13 jours et en reçoit plus ou moins 6 %. Le vent, l'humidité ambiante et la température, qui agissent sur la viabilité du pollen, sont des facteurs à prendre en compte pour comprendre les possibilités de croisement entre espèces ou variétés.
La figure 3 illustre ce qui se passe lorsque la fleur de l'épillet émet son pollen à l'extérieur (les 5 à 10 %), ce qui donne une chance à l'allogamie (échange génétique) et prouve que, dans la nature, le blé a besoin de se régénérer à l'extérieur de temps en temps.
La Polyploïdisation : Un "Accident" Génétique Constructif
Outre ce croisement déjà rare pour le blé, la démultiplication du génome est encore plus inhabituelle, mais a été constatée plusieurs fois par les généticiens dans le groupe des graminées auquel le blé appartient. Ainsi, une graminée sauvage, une égilope, qui possède une paire de chromosomes BB, a pu polliniser le blé Urartu, un cousin de l'engrain aux chromosomes AA, et donner par fusion des deux paires de chromosomes, l'amidonnier. Ce dernier comporte ainsi deux paires de 7 chromosomes, AA + BB, et se trouve toujours à l'état sauvage.
Le terme d'accident génétique est parfois employé pour expliquer qu'ici, il ne s'agit pas de croisement entre deux génomes, mais d'une duplication et superposition d'un génome entier sur un autre, dénommée polyploïdisation. Cela explique pourquoi certains pro-OGM osent avancer que le blé serait en fait le premier OGM. Sauf que dans ce cas, la barrière des genres, et même parfois des espèces, n'est pas transgressée.
L'Évolution du Blé : De l'État Sauvage à la Culture Sédentaire
Le passage de l'état dit sauvage ou naturel, non cultivé et exploité par le monde nomade de cueilleurs-récolteurs, à l'état de culture entretenue par un monde sédentarisé, se serait opéré sur dix millénaires selon Kent V. Flannery. L'humanité a eu le temps de s'adapter. C'est dans le Croissant fertile, et assez souvent dans le massif des Zagros, que l'on a actuellement découvert les plus anciennes traces d'établissement de culture (9000 ans avant J.-C.), bien que cela soit contestable et sujet à révision selon les futures découvertes archéologiques. Des fouilles récentes en Israël et en Jordanie ont fait évoluer les connaissances.
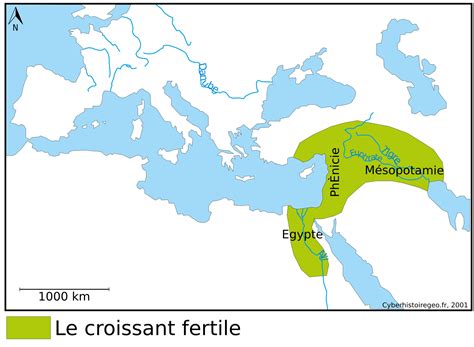
Les Premières Traces de l'Engrain et de l'Amidonnier Cultivés
D'après les dernières recherches, les plus anciens recensements de l'engrain cultivé, au rachis non fragile, ont été réalisés dans les fouilles :
- d'Ali Kosh (Iran-Khuzistan) de 9500 à 8750 avant J.-C.
- Jarmo (Irak-Kurdistan) en 8700 avant J.-C.
- Cayonu Teperi (Sud-Est de la Turquie) en 9000 avant J.-C.
- Abu Hureyra (Nord Syrie) en 9000 avant J.-C.
- Hacilar (Centre-Est de l'Anatolie turque) de 8900 à 8600 avant J.-C.
Pour l'amidonnier cultivé, on en a identifié dans les fouilles de Ramad (Sud-Ouest de la Syrie) en 8200 et 8000 avant J.-C., et de Can Hassan (Sud de l'Anatolie) en 8900 et 8600 avant J.-C.
On observe que cette autogamie du blé signifie qu'il ne se croise pas et ne se modifie pas facilement. En milieu naturel et sans intervention humaine, une fécondation par des gènes provenant d'autres variétés est considérée comme occasionnelle et rare. Cependant, c'est sur la base de cette rareté ou de cet "accident" que le froment que nous connaissons aujourd'hui existe. De l'état sauvage à l'état cultivé, des croisements et des polyploïdisations d'espèces se sont opérés durant les millénaires de la domestication du blé.
L'Arbre Généalogique du Froment : Des Disciplines Scientifiques Récentes
L'étude morphologique (de l'apparence extérieure), cytogénétique (des chromosomes) et moléculaire du blé permet d'esquisser l'évolution ou l'arbre généalogique du froment. Ces disciplines scientifiques récentes sont essentielles pour des définitions plus précises de la généalogie du blé. Ce parcours généalogique n'a pu être réalisé que près de 8 000 ans plus tard. L'importance des chromosomes dans l'hérédité a été mise en lumière dans les années 1920 par l'Américain Thomas H. Morgan, qui a obtenu le prix Nobel de physiologie et de médecine en 1933 pour ses travaux. En 1949, la première datation au carbone 14 de deux échantillons de bois provenant de tombes égyptiennes a été réalisée par Willard Frank Libby (prix Nobel de chimie en 1960). Le terme "palynologie", forgé en 1944, est aujourd'hui universellement employé pour désigner l'ensemble des recherches archéologiques sur les graines et leurs pollens conservés dans les couches terrestres inférieures. La relative "jeunesse" de ces connaissances scientifiques explique qu'elles aient parfois du mal à s'imposer face à ce qu'il est convenu d'appeler la connaissance de terrain.
La Classification Traditionnelle du Blé : Une Connaissance de Terrain
Sur le terrain, ce ne sont bien sûr pas les chromosomes, invisibles à l'œil nu, qui permettent de différencier les blés. Les agriculteurs classent plus volontiers en parlant de blés nus ou vêtus, grains durs ou tendres, barbus ou non-barbus, des repères tangibles et visuels. C'est ainsi que tous les grains vêtus sont parfois appelés « épeautre » en français, ou « farro » en italien. Ces 8 000 années autorisent parfois le paysan en Provence à appeler l'engrain « petit épeautre », l'amidonnier sera l'« épeautre de Tartarie » et, par conséquent, l'épeautre (spelta) en devient le « grand épeautre ».
Les Caractéristiques des Céréales Sauvages : Une Adaptation à l'Auto-Ensemencement
Les premières céréales, à l'état naturel dites « sauvages », sont d'abord cueillies plutôt que récoltées par les peuples nomades. Elles ont alors une physionomie adaptée à l'auto-ensemencement. Il est aujourd'hui possible de reconnaître une céréale sauvage à plusieurs points distinctifs.

Le Rachis Fragile et la Dissémination des Graines
Tout d'abord, le rachis, axe ou colonne centrale où sont disposés en arborescence les épillets, est fragile. Cette fragilité permet aux graines, une fois à maturité, de se séparer, une à une, à l'entre-nœud de l'épi et ainsi de se disséminer au vent. Ce processus est appelé égrenage ou semis spontané. La forme aérodynamique des barbes de la graine influence son comportement lors de sa chute. Cette barbe n'est pas seulement une défense naturelle, c'est aussi un dispositif naturel qui permet aux graines dispersées par le vent de bien se planter dans le sol.
Les Mérithalles : Un Dispositif d'Ancrage Naturel
Enfin, et c'est ce qui différencie le grain sauvage du grain cultivé, il dispose à sa base, au bas de la glume, d'espèces de cils coriaces et hérissés installés à rebrousse-poil (appelés mérithalles) qui lui permettent de s'accrocher et s'incruster dans la terre meuble, sans pouvoir en ressortir. Les graines étant une source de nourriture pour de nombreux animaux, en particulier les oiseaux, ces caractéristiques décrites précédemment étaient essentielles pour leur défense et leur reproduction.
La Domestication du Blé et les Premières Sélections Humaines
Les humains ont progressivement choisi de s'établir et de cultiver. Ainsi, une grande partie de l'humanité est passée de la vie nomade à la vie sédentaire. Cependant, cette perspective doit être nuancée en lisant l'éclairage de David Graeber, car ce passage n'a pas été aussi net et aussi choisi que l'histoire abrégée le mentionne parfois.
Heureusement pour ces femmes et ces hommes qui ont choisi de le domestiquer, le blé sauvage donnait de temps à autre des grains que nous qualifierons ici d'anormaux. Ils ne seront considérés comme anormaux que temporairement, car par la suite, ils deviendront la norme. Ces grains seront préférés et sélectionnés par les premiers cultivateurs. En effet, la séparation ou l'éclatement des épillets de l'épi une fois à maturité ne permettant pas une récolte facile, l'anomalie pour les épillets consistait à ne pas se disperser au vent une fois mûrs. Il n'était plus nécessaire d'aller les ramasser par terre aux alentours. On pouvait les récolter sur l'épi, puis les battre à façon, en fonction des besoins. Cependant, il fallait ensuite obligatoirement les réensemencer, donc les cultiver. Toutes ces métamorphoses représentent, en somme, les premières sélections du blé. Bien d'autres sélections suivront.
L'Intéressante Qualité Originelle du Blé
Une étude menée par l'Université du Saskatchewan au Canada s'est penchée de manière approfondie sur les caractéristiques des ancêtres du froment panifiable. Les propriétés des variétés originelles peuvent surprendre. Si une augmentation des fibres dans la farine ne surprend pas, compte tenu du petit format du grain, le pourcentage de protéines, passant de 11,5 % à presque 20 %, est remarquable. Bien que ces protéines ne soient pas forcément de qualité "machinable", elles contribuent à une meilleure qualité nutritionnelle grâce à des teneurs en acides aminés de meilleure valeur nutritionnelle. La teneur en acides gras, sels minéraux essentiels et vitamines est également supérieure à celle de nos blés actuels. Ensuite viennent les éléments antinutritionnels…