La cellule est l'unité fondamentale, structurale et fonctionnelle des organismes vivants, une entité vivante qui fonctionne de manière autonome, tout en restant coordonnée avec les autres. De taille variable, chaque cellule possède toutefois la même structure et communique entre elles, remplissant toutes les fonctions de l’organisme, à savoir le métabolisme, le mouvement, la croissance, la reproduction ou encore la transmission de gènes. Comprendre sa structure et ses interactions est essentiel pour appréhender les processus biologiques complexes.
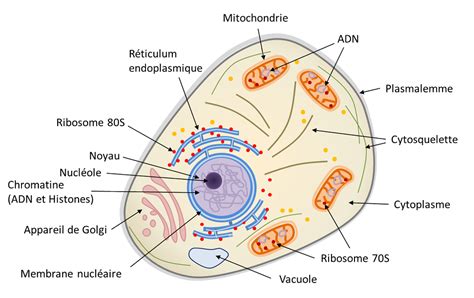
I. L'Architecture Cellulaire : Une Unité Fondamentale
Chaque cellule est délimitée par une membrane plasmique et contient un cytoplasme où résident divers organites spécialisés.
1. La Membrane Plasmique : La Porte d'Entrée et de Sortie de la Cellule
La membrane plasmique ou cellulaire est composée de phospholipides qui lui confèrent sa structure de base, auxquels s’ajoutent des molécules de cholestérol qui augmentent l’imperméabilité et la rigidité de la membrane. Elle est également composée de protéines qui assurent la plupart de ses fonctions spécifiques. Par le caractère hydrophobe de sa couche lipidique, la membrane plasmique de la cellule ne permet pas le passage de molécules polaires (électrolytes) et de macromolécules. Celle-ci est cependant semi-perméable, car la cellule doit réguler ses concentrations ioniques.
1.1. Les Échanges Transmembranaires : Maintenir l'Équilibre Cellulaire
La cellule se nourrit en puisant dans le milieu extracellulaire les nutriments nécessaires au maintien de son activité et de sa croissance. Elle rejette dans ce même milieu les produits de dégradation inutiles ou toxiques. L’ensemble des échanges vont ainsi se faire à travers la membrane plasmique ou à l’aide de protéines transmembranaires.
Osmose : La Diffusion de l'EauC’est le processus de la diffusion de l’eau dans différents milieux. Si le liquide extracellulaire est moins concentré que la cellule, on dit que le milieu est hypotonique. Si le liquide extracellulaire est plus concentré que la cellule, on dit que le milieu est hypertonique.
Transport Actif : Contre le Gradient de ConcentrationLe transport actif exige de l’énergie (ATP) pour rendre la structure transporteuse capable de fonctionner contre un gradient de concentration. Il est utilisé principalement pour le transport des macromolécules (ex : glucose, acides aminés,…) ou des électrolytes (ex : sodium, potassium…). Un exemple notable est la pompe Na+/K+/ATP. La concentration en sodium (Na+) est supérieure à l'extérieur de la cellule, tandis que la concentration en potassium (K+) est supérieure dans le milieu intracellulaire.
Endocytose : La Capture CellulaireÀ l'échelle cellulaire, on parle d’endocytose lorsque la cellule capture des éléments, par invagination de sa membrane plasmique, pour former des vacuoles dans son cytoplasme.
Protéines Transmembranaires : Des Portes SpécifiquesCertains transports nécessitent l’intervention de protéines présentes dans la membrane cellulaire. Ces protéines transpercent la membrane de part en part et fonctionnent comme des portes, régulant le passage de substances.
2. Le Cytoplasme : Le Cœur Actif de la Cellule
Le cytoplasme désigne le contenu d'une cellule vivante, englobant le cytosol et les organites.
2.1. Le Cytosol et le Cytosquelette : Structure et Mouvement
Le milieu intracellulaire se compose d'un liquide appelé cytosol. Il est constitué en moyenne de 85 % d'eau et son pH est neutre. Ce réseau fibreux de nature protéique constitue à la fois un squelette et une musculature pour les cellules. Le cytosquelette contient les centrosomes, des éléments tubulaires intervenant dans la division cellulaire. Chaque centrosome est composé de deux centrioles, ce sont des structures cellulaires intracytoplasmiques constituées de 9 triplets de 3 tubules. Chaque cellule contient donc deux centrioles (perpendiculaires et ne se touchant pas) et l'ensemble forme le centrosome qui est toujours à proximité du noyau.
2.2. Les Organites Spécialisés : Des Fonctions Clés pour la Vie Cellulaire
Le cytoplasme abrite une multitude d'organites, chacun avec des rôles spécifiques.
Réticulums Endoplasmiques (RE) : Synthèse et MaturationLes réticulums endoplasmiques (RE) sont des organites avec une double membrane intracellulaire et ressemblent à un amas de replis formant des cavités, appelées "citernes". Ils sont en continuité avec la membrane du noyau. Le réticulum endoplasmique granuleux (REG) ou réticulum endoplasmique rugueux (RER) a sa surface recouverte de ribosomes qui assemblent les acides aminés en protéines suivant l'information venue du noyau. Le réticulum endoplasmique lisse (REL), lui, n'en porte pas.
Ribosomes : Les Usines à ProtéinesCes sphères, libres ou associées au REG, participent à la synthèse protéique à partir d'ARN (traduction). Leur fonction est de synthétiser les molécules de protéines à partir des acides aminés.
Appareil de Golgi : Le Centre de Tri et de SécrétionIl est formé de sacs aplatis les uns sur les autres. Son rôle est de stocker les protéines issues du REG, d'achever leur maturation, de vérifier leur qualité et de les sécréter. Les protéines à sécréter sont alors concentrées dans des vésicules issues des extrémités de l'appareil de Golgi.
Lysosomes : Les Recyclers CellulairesCe sont des vésicules contenant des enzymes hydrolytiques qui proviennent du RE ou de l'appareil de Golgi. Ces enzymes servent à digérer les macromolécules inutilisables telles que les organites détruits ou abimés, les substances toxiques.
Mitochondries : Les Centrales ÉnergétiquesLes mitochondries sont des organites en forme de haricot de très petite taille, ressemblant à de petits bâtonnets et possédant une double membrane intracellulaire. La membrane interne forme des replis appelés crêtes, qui s'imbriquent dans une substance appelée matrice. On y trouve de nombreuses enzymes responsables de la dégradation des nutriments sous forme simple, tel le glucose. Ces dégradations se déroulent en présence d'oxygène et portent le nom de respiration cellulaire. Elles permettent aux mitochondries de former de l'ATP, source d'énergie de la cellule. Le nombre de mitochondries d'une cellule dépend de l'intensité de son activité : une cellule musculaire, par exemple, en possède beaucoup. Ces organites possèdent leur propre matériel génétique, l’ADN mitochondrial, hérité par la mère.
3. Le Noyau : Le Centre de Contrôle Génétique
Le noyau, le plus gros des organites, est visible dans la cellule lorsqu'elle ne se divise pas (interphase). Il possède un diamètre variant de 10 à 20 µm et est entouré par une double membrane : la membrane nucléaire. Cette membrane nucléaire contient des pores permettant les échanges nucléocytoplasmiques dans les deux sens. Le nucléoplasme est le liquide dans lequel baignent les éléments contenus dans le noyau. C'est là que l'ADN, le matériel génétique, est organisé en chromosomes.
II. La Communication Cellulaire : Un Dialogue Essentiel
La vie multicellulaire repose sur un ensemble complexe d'interactions entre les cellules elles-mêmes, ainsi qu’avec le milieu extracellulaire.
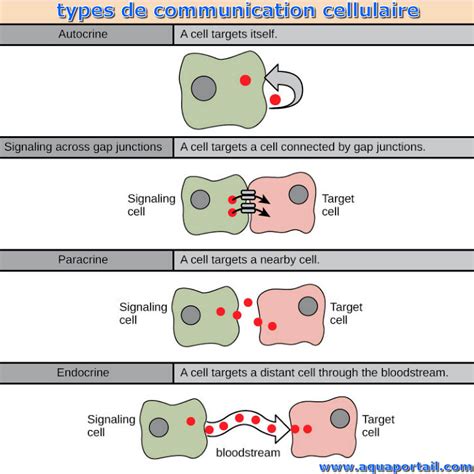
1. La Communication à Distance : Messagers et Récepteurs
Pour communiquer entre elles, les cellules éloignées synthétisent des messagers (enzymes, médiateurs, hormones…) ainsi que des récepteurs.
1.1. Les Molécules Messagères : Porteurs d'Information
Une molécule libérée par une cellule A est informative lorsqu'elle interagit spécifiquement avec une structure appelée récepteur d'une cellule B pour y initier des réactions conduisant à des effets spécifiques. Les messagers sont de nature chimique diverse : dérivés d'acides aminés (noradrénaline, angiotensine…), des dérivés d'acides gras (prostaglandines…) ou des dérivés de cholestérol (cortisol, stéroïdes…). Leur classification repose sur la distance qui sépare leur site de libération de leur site d'action.
Hormones : Le Système EndocrineLes hormones sont véhiculées par le sang depuis la glande qui les libère jusqu'à l'organe où elles exercent leurs effets, agissant sur des cellules cibles éloignées.
Médiateurs : La Transmission NerveuseLes médiateurs sont libérés à l'extrémité d'un nerf, ils transmettent une information à une structure qui peut être un nerf ou un muscle, facilitant la communication rapide dans le système nerveux.
1.2. Les Récepteurs : Les Décodeurs de Messages
Un récepteur est une structure moléculaire de nature polypeptidique interagissant avec un messager. Cette interaction crée alors une modification du récepteur induisant un effet biologique. Les récepteurs peuvent être membranaires (absence de pénétration du médiateur dans la cellule) ou intranucléaires (pénétration du médiateur dans la cellule).
- Récepteurs Membranaires : Interaction en SurfaceUn récepteur membranaire comporte une partie extracellulaire, où se trouve le site de reconnaissance de la molécule informative, une partie transmembranaire et une partie intracellulaire. L'activation du récepteur membranaire ne nécessite pas la pénétration de la molécule informative dans la cellule. Cette activation va induire des modifications qui vont soit être localisées à la membrane, soit s'étendre au cytoplasme ou bien atteindre le noyau. Parmi eux, on distingue :
- Récepteurs Canaux : Régulateurs d'IonsIls comportent un canal faisant communiquer le cytoplasme avec le milieu extracellulaire. Le messager module l'ouverture du canal et régule l'entrée dans la cellule des ions (Na+, K+, Ca2+, Cl- ;..).
- Récepteurs Liés aux Protéines G : Cascade de SignalisationLeur activité nécessite la présence de GDP (guanosine diphosphate) pour initier une cascade de signalisation intracellulaire.
- Récepteurs Enzymes : Catalyse DirecteLe récepteur possède sa propre activité enzymatique, permettant une réponse directe après liaison du messager.
2. La Communication par Contact Direct : Interactions Cellule-Cellule
Les cellules proches interagissent directement par contact. Elles vont pouvoir échanger des informations en adhérant l'une à l'autre à l’aide de jonctions cellulaires.
Desmosomes : Ancrages CellulairesLes desmosomes sont composés de plaques circulaires cytoplasmiques denses, situées à l’intérieur de chaque cellule, offrant une forte adhérence.
Jonctions Communicantes : Canaux d'Échange DirectCes jonctions sont formées de connexines, des protéines membranaires qui, lorsqu’elles s’associent, forment une connexon, permettant un échange direct de petites molécules entre cellules.
III. Le Repiquage Cellulaire : Une Technique Fondamentale en Biologie
Le repiquage cellulaire est une technique cruciale en culture cellulaire, permettant de maintenir la viabilité et la prolifération des cellules en laboratoire.
1. La Réplication de l'ADN et la Division Cellulaire : Le Fondement du Repiquage
L'ADN peut se répliquer, grâce à une ADN polymérase et grâce à l'ouverture du double brin. Chaque brin de la molécule « mère » sert de matrice pour la synthèse d'un nouveau brin. La synthèse se fait dans le sens 5' vers 3' et face à chaque nucléotide, la polymérase accroche au brin en cours de synthèse le nucléotide complémentaire suivant la règle A s'associe à T (et réciproquement) et G s'associe à C (et réciproquement). À l'issue de la réplication, à partir d'une molécule d'ADN double brin, on se retrouve avec deux molécules d'ADN filles double brin, chacune identique à la molécule mère de départ. Chacune des deux nouvelles molécules obtenues est formée d'un brin de la molécule d'origine et d'un nouveau brin synthétisé à partir des nucléotides incorporés. On dit que la réplication de l'ADN est semi-conservative. Au niveau cellulaire, ce phénomène de réplication de l'ADN est couplé à la division cellulaire, à l'issue de laquelle on va obtenir deux cellules filles qui auront le même patrimoine génétique que celui de la cellule mère dont elles sont issues.
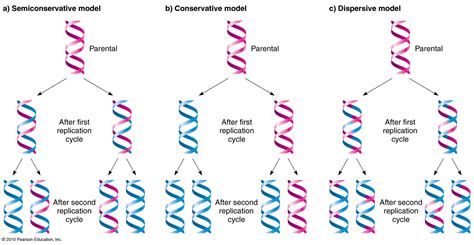
L'ADN est réparti entre différents chromosomes. Quand une cellule se divise, les chromosomes se condensent et passent d'une structure relâchée à une structure condensée. Dans la structure relâchée, les chromosomes possèdent une seule chromatide (1 ADN double brin), dans la structure condensée, ils possèdent deux chromatides (identiques) reliées par le centromère. Elle permet la répartition des chromosomes en deux lots identiques.
2. La Culture Cellulaire : Un Environnement Contrôlé
La culture cellulaire présente de nombreux avantages mais aussi des inconvénients. Les cellules sont dans un environnement contrôlé (température, nutriments disponibles, éventuellement en présence de matrice extracellulaire…) mais cet environnement est parfois éloigné des conditions physiologiques. Dans des cultures classiques en 2D, chaque cellule est facilement visible, mais les propriétés des organisations 3D dans les tissus peuvent être perdues.
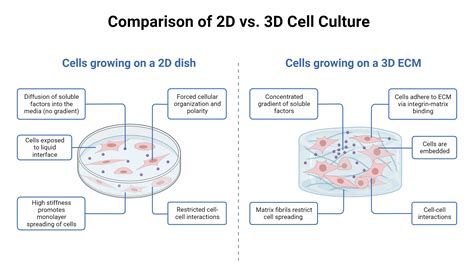
Les cellules sont cultivées sur du plastique traité pour que les cellules puissent y adhérer ou sur des éléments de matrice extracellulaire que l’on dépose avant d’ensemencer les cellules (collagène, fibronectine ou encore le Matrigel (lame basale produite par des cellules de sarcome de souris) qui, par son épaisseur, permet de créer un environnement 3D). Les cellules sont cultivées dans un milieu nutritif contenant des nutriments et des vitamines et souvent additionné de sérum (en général du sérum de veau fœtal) dans lequel se trouvent des facteurs de croissance nécessaires à la prolifération.
2.1. Suivi de la Confluence et de l'Adhérence
Pour les cellules qui prolifèrent, on suit le niveau de confluence, c’est-à-dire le pourcentage de la surface couvert par les cellules. Le suivi en temps réel de l’adhérence et de la confluence des cellules peut être réalisé par la technologie xCELLigence, qui est basée sur les variations de l’impédance (de la résistance électrique) d’une couche riche en or qui dépend des cellules qui sont attachées sur le substrat. Ces modifications d’impédance sont traduites en un « cell index » qui est nul si aucune cellule n’est attachée. Des changements dans la morphologie cellulaire (adhérence, étalement), le nombre de cellules (prolifération ou mort) ou la migration peuvent être étudiés à l’aide de ce système. On suit l’adhérence initiale de cellules que l’on vient de disperser dans le milieu de culture. À plus long terme, on peut suivre l’étalement des cellules. Ici, les cellules ne prolifèrent pas, mais si elles proliféraient on observerait une pente ascendante au lieu d’un plateau.
2.2. Le Processus de Repiquage
Le passage des cellules confluentes en culture cellulaire consiste à détacher, à diluer puis à redéposer des cellules dont la densité avait atteint le maximum autorisé sur la surface de culture. Lorsque les cellules deviennent confluentes, leur prolifération ralentit en raison du contact cellulaire et du manque d’espace. Pour maintenir leur état physiologique et leur capacité de division, on procède à une dissociation mécanique et/ou enzymatique (souvent à la trypsine-EDTA (la trypsine digère la matrice extracellulaire et l’EDTA piège les ions Ca2+ nécessaires aux cadhérines qui permettent l’adhérence des cellules entre elles)), suivie d’un comptage et d’un repiquage dans de nouveaux flacons à une densité adaptée.
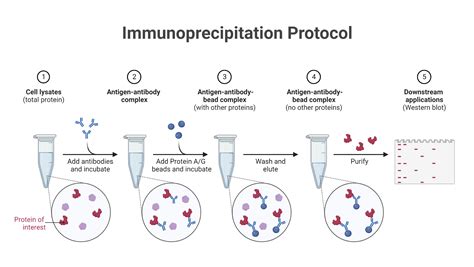
Protocole pour la Culture Cellulaire
3. Les Lignées Cellulaires Immortelles : Un Outil Précieux
Les cellules HeLa, isolées en 1951 à partir d’un carcinome du col de l’utérus d’Henrietta Lacks (sans son consentement), ont constitué la première lignée cellulaire humaine immortelle que les chercheurs ont pu faire se diviser indéfiniment in vitro. Malgré les problèmes éthiques liés à leur prélèvement, elles demeurent un outil fondamental en biologie cellulaire, apparaissant dans plus de 120 000 publications scientifiques. Les lignées de cellules cancéreuses ne connaissent pas ces phénomènes de sénescence, contrairement aux cellules normales qui, après un certain nombre de passages, entrent en sénescence, un état de non-prolifération irréversible. Un test de sénescence dans des cultures de fibroblastes de souris embryonnaires (MEF) ou test SABG (Senescence-Associated Beta-Galactosidase) montre qu'avant la sénescence, les MEF au 2ème passage après prélèvement ont une forme de fuseau. Après le 8ème passage, les MEF se sont agrandis et se sont aplatis. La coloration bleu-vert indique l’expression de la β-galactosidase associée à la sénescence (la coloration est obtenue en présence du substrat X-gal).
IV. Techniques d'Étude des Protéines Cellulaires : Allant au-delà du Repiquage
Au-delà du simple repiquage, de nombreuses techniques permettent d'analyser les composants cellulaires, en particulier les protéines.
1. Le Fractionnement Cellulaire : Isoler les Compartiments
Le fractionnement cellulaire est une technique pour purifier des fractions particulières du contenu des cellules (noyau, mitochondries, ribosomes…). Dans une étude des fractions protéiques cytoplasmiques et nucléaires dans un modèle de maladie de Huntington, on soumet des neurones avec des allèles sauvages (+) ou mutants (109) pour le gène codant la huntingtine à un fractionnement cellulaire par centrifugation qui permet de récupérer les noyaux dans le culot et avec le cytoplasme qui se retrouve dans le surnageant. On réalise des extraits protéiques de ces deux fractions que l’on analyse ensuite par western-blots avec des anticorps reconnaissant la protéine REST/NRSF ainsi que des anticorps reconnaissant la tubuline (contrôle des fractions cytoplasmiques) et Histone H1 (contrôle des fractions nucléaires). On constate que dans les neurones mutants, REST/NRSF a une localisation nettement plus nucléaire que dans les neurones sauvages au détriment de la localisation cytoplasmique. Le fractionnement a été correctement réalisé car on ne retrouve pas de tubuline dans les fractions nucléaires et pas d’Histone H1 dans les fractions cytoplasmiques.
2. Le Western Blot : Détecter et Quantifier les Protéines
Le western-blot est une technique couramment utilisée pour détecter et quantifier des protéines spécifiques dans un échantillon. Les cellules sont lysées dans du tampon RIPA (150 mM NaCl, 1 % NP-40, 0,25 % Na-désoxycholate, 50 mM Tris-HCl [pH 7,4]) contenant 1 comprimé d’inhibiteur de protéase et 1 comprimé d’inhibiteur de phosphatase par 10 ml. La concentration totale de protéines dans les lysats est déterminée à l’aide d’un kit Pierce BCA Protein Assay. L’absorbance est mesurée à l’aide d’un spectrophotomètre à microplaque. Un gel de polyacrylamide à 8 % contenant du SDS surmonté d’un gel de stacking (pour concentrer les protéines) est préparé. Ensuite, 15 µL d’échantillon, contenant 9 µg de protéines totales et 5 µL de tampon d’échantillon (5,7 ml d’eau, 1,6 ml de glycérol (densité importante pour que l’échantillon aille au fond du puits), 1,1 ml de SDS à 10 % (dénaturation), 1,3 ml de Tris 0,5 M (pH 6,8), 25 mg de dithiotréitol (DTT, casse les ponts disulfures des protéines), 300 µL de bleu de bromophénol (pour bien visualiser l’échantillon)) sont chauffés à 90 °C pendant 5 min (pour dénaturer les protéines), refroidis sur de la glace et chargés sur le gel. Le gel est incubé dans du tampon d’électrophorèse (Tris Base 25 mM, glycine 190 mM, 0,1 % SDS) à 70 V jusqu’à ce que les échantillons atteignent le gel de séparation, puis à 150 V jusqu’à ce que les échantillons atteignent le bas du gel. Ensuite, les protéines sont transférées sur une membrane en polyfluorure de vinylidène (PVDF) pendant 1 h à 80 V sur glace dans un tampon de transfert froid (Tris base 25 mM, glycine 190 mM, éthanol 20%). La membrane est rincée à l’eau et lavée 2x dans une solution saline tamponnée au Tris et du Tween-20 (TBS-T) (20 mM de Tris/HCl, 137 mM de NaCl, 0,1 % de Tween-20). La membrane est incubée pendant 1 h à température ambiante dans un tampon de blocage (5% BSA dans du TBS-T, sature les sites non spécifiques sur lesquels l’anticorps pourrait s’accrocher) sous agitation. Ensuite, la membrane est incubée pendant la nuit dans le tampon de blocage avec un anticorps primaire à la concentration souhaitée à 4°C sous agitation. La membrane est lavée 3 × 5 min dans du TBS-T et incubée dans le tampon de blocage avec l’anticorps secondaire reconnaissant l’anticorps primaire pendant 1 h à température ambiante. Un ECL est réalisé pour la détection. Une membrane de western-blot colorée au Rouge Ponceau colore en rouge l’ensemble des protéines qui ont été transférées depuis le gel de polyacrylamide vers la membrane (de nitrocellulose ou de PVDF). Cela permet de contrôler si le transfert s’est bien passé et si la quantité totale des protéines est similaire d’un puits à l’autre (ici, il y a un peu moins de protéines dans le puits à gauche).
3. L'Immunoprécipitation : Isoler des Complexes Protéiques
Cette technique permet d’isoler une protéine d’un extrait cellulaire et tous les interactants directs ou indirects avec cette protéine grâce à l’interaction très spécifique anticorps-antigène. Les anticorps peuvent être fixés sur des billes et on peut récupérer dans le culot les complexes formés après une centrifugation. Alternativement, on peut utiliser des billes métalliques qui sont ensuite attirées par un aimant. Cette technique permet de savoir si deux protéines A et B appartiennent à un même complexe à un moment donné et dans des conditions données. On fait une immunoprécipitation avec un anticorps contre A et on analyse la présence par western-blot de la protéine B reconnue par un autre anticorps.
Un exemple d’immunoprécipitation est la mise en évidence de la présence dans un même complexe des protéines BRCA1 et ER-α. Des extraits protéiques de cellules de tumeurs de glande mammaire MCF-7 (qui expriment beaucoup de récepteurs aux œstrogènes) et de tumeurs de prostate Du-145 (qui expriment très peu de récepteurs aux œstrogènes) sont soumis à une immunoprécipitation (IP) avec un anticorps anti-BRCA1 ou anti-ER-α. Une immunoprécipitation avec un anticorps Ig est réalisée à titre de témoin négatif (puits à gauche). Ces fractions immunoprécipitées sont soumises à un western-blot avec un anticorps anti-BRCA1 ou anti-ER-α. On vérifie d’abord qu’il y a bien du ER-α et du BRCA1 dans la fraction précipitée avec leurs propres anticorps respectifs. On constate ensuite la présence de ER-α dans la fraction précipitée avec l’anticorps anti-BRCA1 et la présence de BRCA1 dans la fraction précipitée avec l’anticorps anti-ER-α, seulement dans les cellules MCF-7.
L'interaction entre Myc-MT1X et HA-FHL3 peut également être démontrée par immunoprécipitation. On ne dispose pas de bons anticorps reconnaissant spécifiquement la protéine MT1X et FHL3. Des cellules 293T ont été transfectées avec des plasmides permettant d’exprimer des protéines MT1X couplées au tag Myc et FHL3 couplées au tag HA. Des extraits protéiques sont réalisés à partir de ces cellules et traitées en western-blot avec des anticorps anti-HA et anti-Myc (input). On réalise ensuite une immunoprécipitation (IP) avec un anticorps IgG (témoin négatif) et un anticorps anti-HA (ce qui revient à immunoprécipiter HA-FHL3) et on réalise ensuite une analyse par western-blot du culot obtenu avec un anticorps anti-HA (on vérifie que l’immunoprécipitation a bien fonctionné) et avec un anticorps anti-Myc (on cherche à savoir si Myc-MT1X se trouve dans le culot avec HA-FHL3 montrant que FHL3 et MT1X sont présents dans un même complexe).
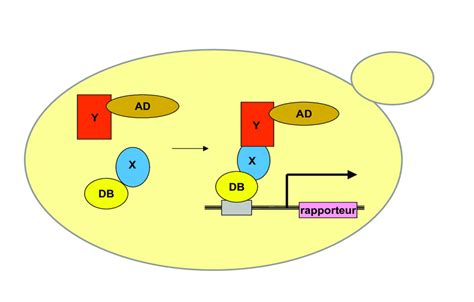
4. Le Système du Double Hybride : Révéler les Interactions Protéine-Protéine
Le système du double hybride effectué chez la levure ou dans des cellules de mammifères permet de mettre en évidence l’interaction entre deux protéines. Il est basé sur la nature modulaire des facteurs de transcription qui sont généralement composés de deux domaines : un domaine de liaison à l’ADN et un domaine d’activation de la transcription. Chez la levure, on utilise le facteur de transcription Gal4. Si on veut savoir si la protéine A interagit avec la protéine B, on fait synthétiser aux cellules deux protéines de fusion : une avec le domaine de liaison à l’ADN de Gal4 fusionné avec la protéine A et une autre avec le domaine d’activation de Gal4 fusionné avec la protéine B. Les protéines fusion A-Gal4DBD (DBD = domaine de fixation à l’ADN) et B-Gal4AD (AD = domaine d’activation de la transcription) doivent interagir pour que la transcription du gène rapporteur (en orange) puisse être activée. Le domaine de fixation à l’ADN de Gal4 se fixe sur une séquence appelée UAS (ne pas tenir compte du Gal1).
Un exemple d’analyse en double-hybride avec la levure montre la croissance de colonies de levure sur milieu avec histidine (+His) ou sans histidine (-His). Les levures sont déficientes en l’enzyme HIS3 qui permet de synthétiser l’histidine qui est indispensable pour la croissance mais elles possèdent une construction où le gène codant cette enzyme est sous le contrôle d’une séquence UAS. Les levures exprimant la protéine DDB1 fusionnée au domaine de liaison à l’ADN de GAL4 (BD-DDB1) sont co-transformées soit avec le vecteur AD vide (juste le domaine d’activation de la transcription de GAL4), soit avec les constructions AD-CUL4 ou AD-DET1 où ce domaine est fusionné avec les protéines CUL4 ou DET1, respectivement. Différentes doses de plasmides avec les différentes constructions AD sont testées (gradients horizontaux en haut). En présence d’histidine (+His), toutes les souches poussent, montrant qu’il n’y a pas de toxicité liée à la transformation. En absence d’histidine (−His), seule la co-expression BD-DDB1/AD-CUL4 et BD-DDB1/AD-DET1 permet la croissance, indiquant l’activation du gène rapporteur HIS3 par interaction spécifique entre DDB1 et CUL4 ou DET1. On remarque un effet dose plus marqué pour AD-DET1 montrant que l’interaction entre DDB1 et DET1 est plus fragile qu’entre DDB1 et CUL4.
5. Immunochimie et Immunofluorescence : Visualiser les Protéines in Situ
Comme le western-blot, ces méthodes utilisent la spécificité de reconnaissance des anticorps pour mettre en évidence les protéines d’intérêt mais cette fois-ci non pas à partir d’extraits protéiques mais dans des cellules (cyto) ou des tissus (histo) qui ont été fixés au préalable.
5.1. Immunohistochimie : Localisation Tissulaire
Un exemple d’immunohistochimie sur un embryon de poulet permet de reconnaître les cellules de crêtes neurales (anticorps primaire HNK-1 (en vert) qui reconnaît un antigène à la surface de ces cellules). La fixation est réalisée avec du paraformaldéhyde (PFA) 4% qui donne du formaldéhyde en solution aqueuse. Il permet de préserver les structures cellulaires en pontant les protéines entre elles. On doit réaliser une perméabilisation pour la bonne pénétration des anticorps avec des détergents non ioniques comme le Tween 20 ou le Triton X-100. L’anticorps primaire (vert) reconnaît spécifiquement l’antigène (triangle vert). L’anticorps secondaire (rouge) reconnaît le premier anticorps (il doit être choisi pour reconnaître les anticorps de l’espèce dans laquelle a été produit l’anticorps primaire) et il est couplé à la peroxydase qui, en présence de DAB (3,3′-diaminobenzidine) et d’H2O2, donne un produit coloré brun.
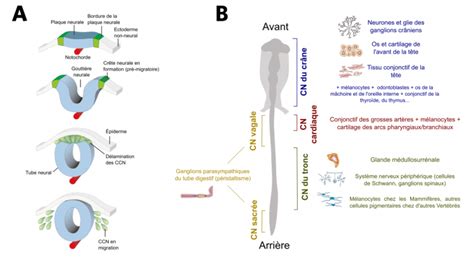
Une immunohistochimie montre que les cellules de crêtes neurales ne migrent que dans la moitié antérieure des somites. Vue latérale de la région troncale d’un embryon de poulet immunomarqué avec l’anticorps HNK-1 qui reconnaît les cellules de crêtes neurales. La partie dorsale est vers le bas. On constate que les cellules de crête neurale, bien qu’elles sortent uniformément le long de l’axe antéro-postérieur du tube neural, migrent uniquement dans la partie antérieure des somites.
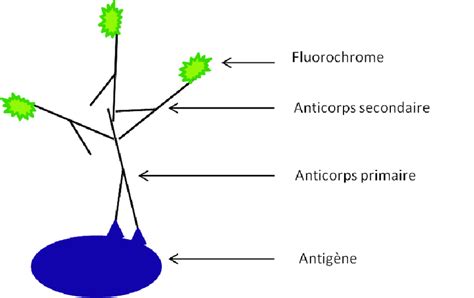
5.2. Immunofluorescence : Détection par Fluorophores
L’immunochimie est basée sur la révélation d’une activité enzymatique apportée par l’anticorps secondaire alors que l’immunofluorescence est basée sur l’émission de photons à une longueur d’onde donnée d’un fluorophore couplé à l’anticorps secondaire. L’anticorps primaire dirigé contre la protéine d’intérêt X est reconnu ensuite par un second anticorps dirigé contre l’anticorps primaire et qui porte un fluorophore, permettant une visualisation précise de la localisation des protéines dans la cellule ou le tissu.
tags: #repiquage #bio #cell #definition